
2 Do Species Change?
bLearning Objectives
- Identify and describe the major developments in scientific thought that led to the discovery of evolutionary processes.
- Explain how natural selection works and results in evolutionary change over time.
- Explain what is meant by the “Modern Synthesis” and its impacts on evolutionary thought.
- Discuss the teaching of human evolution in the U.S. and abroad.
- The Beginnings of Evolutionary Thinking
Throughout our evolutionary history, humans have developed an understanding of the natural world as they interacted with and extracted resources from it. To survive, our earliest ancestors possessed an understanding of the physical environment, including weather patterns, animal behavior, edible and medicinal plants, locations of water, and seasonal cycles. Many ancient cultures, including those of the Americas (Dunbar-Ortiz 2014), Mesopotamia, and Egypt, left writings, hieroglyphics, and stories passed down through oral tradition detailing their understanding of the natural environment, human and zoological anatomy, botany, and medical practices (Moore 1993).
There are also over 2,000 years of organized thought and writing regarding evolution, including contributions from Greek, Roman, and Islamic scholars. Three examples of note are included here. The Greek philosopher Aristotle (384–322 BCE) studied the natural world, publishing several volumes on animals based on systematic observations, rather than attributing what he observed to divine intervention, as his contemporaries were doing (Figure 2.1). Aristotle’s system for the biological classification of nearly 500 species of animals was based on his own observations and dissections, interviews with specialists such as beekeepers and fishermen, and accounts of travelers. His nine book History of Animals, published in the 4th century BC, was one of the first zoological taxonomies ever created. Aristotle’s primary contribution to the classification of biological species was to recognize that natural groups are based on structure, physiology, mode of reproduction, and behavior (Moore 1993, 39).



Aristotle’s History of Animals also placed animals in a hierarchy, ranking animals above plants due to what he claimed were their abilities to sense the world around them and to move. He also graded animals according to their modes of reproduction. Those giving birth to live young were placed above those who laid eggs. Warm-blooded animals ranked above invertebrates. This concept of “higher” and “lower” organisms was expanded upon by scholars in the Medieval period to form the Scala Naturae (Latin for “ladder of being”). This “Great Chain of Being,” depicting a hierarchy of beings with God at the top and minerals at the bottom (Figure 2.2), was thought by medieval Christians to have been decreed by God; in this Great Chain, humans were placed closer to God than other species. Aristotle’s works were rediscovered by Islamic scholars in the ninth century and translated into Arabic, Syriac, Persian, and later into Latin, becoming part of university curriculum in 13th-century Europe (Lindberg 1992), allowing Aristotle’s works and ideas to influence other thinkers for 2,000 years.

Science also owes a debt to many Arabic scholars. One such Islamic scholar and writer, who built upon the ideas of Aristotle, was Abū ʿUthman ʿAmr ibn Baḥr al-Kinānī al-Baṣrī / al-Jāḥiẓ, known as Al-Jahiz (776–868 CE), who authored over 200 books (El-Zaher 2018; Figure 2.3). His most well-known work was the seven-volume Kitab al-Hayawan or Book of Animals, in which he described over 350 species in zoological detail. Importantly, Al-Jahiz introduced the idea and mechanisms of biological evolution 1,000 years before Darwin proposed the concept of natural selection in 1859 (Love 2020). For instance, Al-Jahiz wrote about the struggle for existence, the transformation of species over time, and environmental factors that influence the process, all ideas that were later espoused by western European scientists in the 19th century. Al-Jahiz wrote:
Animals engage in a struggle for existing, and for resources, to avoid being eaten, and to breed. Environmental factors influence organisms to develop new characteristics to ensure survival, thus transforming them into new species. Animals that survive to breed can pass on their successful characteristics to their offspring. [Masood 2009]
Another important early Islamic scientist is Ibn al-Haytham (965–1040), a 10th-century Islamic scholar who contributed a great deal to our understanding of optics and how human vision works (Figure 2.4; Lindberg 1992, 177–180). Born in what is now Iraq, al-Haytham was a scholar of many disciplines, including mathematics, physics, mechanics, astronomy, philosophy, and medicine. He authored some 200 books in his lifetime and was an expert on Aristotle’s natural philosophy, logic, and metaphysics. In relation to evolution, al-Haytham’s methodology of investigation—specifically, using experiments to verify theory—is similar to what later became known as the modern scientific method. He is most famous for discovering the laws of reflection and refraction over 1,000 years ago and inventing the camera obscura, which was incredibly important for the eventual development of photography. His work is credited for its influence on astronomy, mathematics, and optics, inspiring Galileo, Johannes Kepler, and Sir Isaac Newton (Tasci 2020). As an inspirational scientific figure, al-Haytham was celebrated in 2016 by UNESCO as a trailblazer and the founder of modern optics (Figure 2.4). An International Year of Light was named in his honor and a scholarly conference on his contributions was held to coincide with the 1,000th anniversary of the publication of his Kitāb al-Manāẓir (Book of Optics; UNESCO.org 2015).
The writings of these Islamic scholars as well as similar scientific texts from other cultures were unknown to or unacknowledged by Western scientists until recently. Fortunately, many science teachers are now incorporating this content into their classes (Love 2020).
Exercises
Western European Evolutionary Thought
Although there have been many different scientific traditions throughout world history, a new global discourse around science emerged in Western Europe in the 19th century. Europeans pointed to the continuing expansion of their colonial power, as well as their military and technological success, as evidence of the efficacy of Western science, which came to dominate on a global scale (Elshakry 2010). The movement toward a global science centered in Western Europe began with formulation of the Scientific Method.
The Scientific Method was first codified by Francis Bacon (1561–1626), an English politician who was likely influenced by the methods of inquiry established by Ibn al-Haytham centuries prior (Tbakhi and Amr 2007). Bacon has been called the founder of empiricism for proposing a system for weighing the truthfulness of knowledge based solely on inductive reasoning and careful observations of natural phenomena. Ironically, he died as a result of trying to scientifically observe the effects of cold on the putrefaction of meat. On a journey out of London, he purchased a chicken and stuffed it with snow for observation, catching a chill in the process. One week later, he died of bronchitis (Urbach, Quinton, and Lea 2023).
The second important development with regard to evolution was the concept of a species. John Ray (1627–1705), an English parson and naturalist, was the first person to publish a biological definition of species in his Historia Plantarum (History of Plants), a three volume work published in 1686, 1688, and 1704. Ray defined a species as a group of morphologically similar organisms arising from a common ancestor. However, we now define a species as a group of similar organisms capable of producing fertile offspring. In keeping with the scientific method, Ray classified plants according to similarities and differences that emerged from observation. He claimed that any seed from the same plant was the same species, even if it had slightly different traits.
The modern period of biological classification began with the work of Carl von Linne (“Carolus Linnaeus”) (1707–1778), a Swedish scientist who laid the foundations for the modern scheme of taxonomy used today. He established the system of binomial nomenclature, in which a species of animal or plant receives a name consisting of two terms: the first term identifies the genus to which it belongs and the second term identifies the species. His original Systema Naturae, published in 1736, went through several editions. By the tenth edition in 1758, mammals incorporated primates, including apes and humans, and the term Homo sapiens was introduced to signify the latter (Paterlini 2007).
Georges-Louis Leclerc, Comte de Buffon (1707–1788), was a prominent French naturalist whose work influenced prominent scientists in the second half of the 18th century. Buffon’s idea that species change over time became a cornerstone of modern evolutionary theory. His technique of comparing similar structures across different species, called comparative anatomy, is still in use today in the study of evolution. He published 36 volumes of Histoire Naturelle during his lifetime and heavily influenced two prominent French thinkers who were to have significant impacts on our understanding of evolution, Georges Cuvier and Jean-Baptiste Lamarck.

Georges Cuvier (1769–1832) was a paleontologist and comparative anatomist (Figure 2.5). One of his first major contributions to the field of evolution was proof that some species had become extinct through detailed and comprehensive analyses of large fossil quadrupeds (Moore 1993, 111). The idea of extinction was not new, but it was challenging to demonstrate if a fossil species was truly extinct or still had living relatives elsewhere. It was also challenging in that it ran counter to religious beliefs of the time. The Bible’s Book of Genesis was interpreted as saying that all species had been created by God in the seven days it took to create the world and that all created species have survived to this day. Extinction was interpreted as implying imperfection, suggesting God’s work was flawed. Also, given that the Earth was calculated to have been created in 4004 B.C.E., based on biblical genealogies, there would not have been enough time for species to disappear (Moore 1993, 112).
Cuvier was so knowledgeable in this field that he became famous for his ability to reconstruct what an extinct animal looked like from fragmentary remains. He demonstrated that fossil mammoths differed from similar living creatures, such as elephants. His many examples of fossils telling the stories of animals that lived and then disappeared were taken as incontrovertible proof of extinctions (PBS 2001). Where Cuvier went awry was his hypothesis of how extinction worked and its causes. As part of his study of comparative anatomy, Cuvier made observations of stratified layers of rock, or sediment, each containing different species. From this, he drew conclusions that species were “fixed” and did not evolve, but then went extinct, and that different assemblages of fossils occurred at different times in the past, as evidenced by the sedimentary layers (Moore 1993, 118). Cuvier explained this through a theory of catastrophism, which stated that successive catastrophic deluges (akin to Biblical floods) swept over parts of the Earth periodically, exterminating all life. When the waters receded from a particular region, lifeforms from unaffected regions would repopulate the areas that were destroyed, giving rise to a new layer of species that looked different from the layer below it. This theory implied that species were fixed in place and did not evolve and that the Earth was young. In fact, Cuvier postulated that the last catastrophe was a deluge he believed occurred five to six thousand years ago, paving the way for the advent of humans (Moore 1993, 118). Cuvier’s catastrophism became part of an ongoing and vociferous debate between two schools of geology. The catastrophists believed the present state of the earth was the consequence of a series of violent catastrophes of short duration, while the uniformitarians thought it was the result of slow acting geological forces that continue to shape the earth.

James Hutton (1726–1797) was one prominent proponent of uniformitarianism. Based on evidence he found at sites in his native Scotland, Hutton argued that the Earth was much older than previously thought. Examining uniformitarianismthe geology of Siccar Point, a cliff site on the eastern coast of Scotland (Figure 2.6), Hutton concluded that the intersection of the vertical and horizontal rocks represented a gap in time of many millions of years, during which the lower rocks had been deformed and eroded before the upper layers were deposited on top. From this, Hutton argued sediments are deposited primarily in the oceans, where they become strata, or layers of sedimentary rock. Volcanic action uplifts these strata to form mountains, which are then subject to erosion from rain, rivers, and wind, returning sediment to the oceans (Moore 1993, 121). Hutton’s Theory of the Earth (1788) demanded vast periods of time (known as “deep time”) for such slow-working forces to shape the earth. At the time, he was heavily criticized for this view, as it contradicted the biblical version of the history of creation.
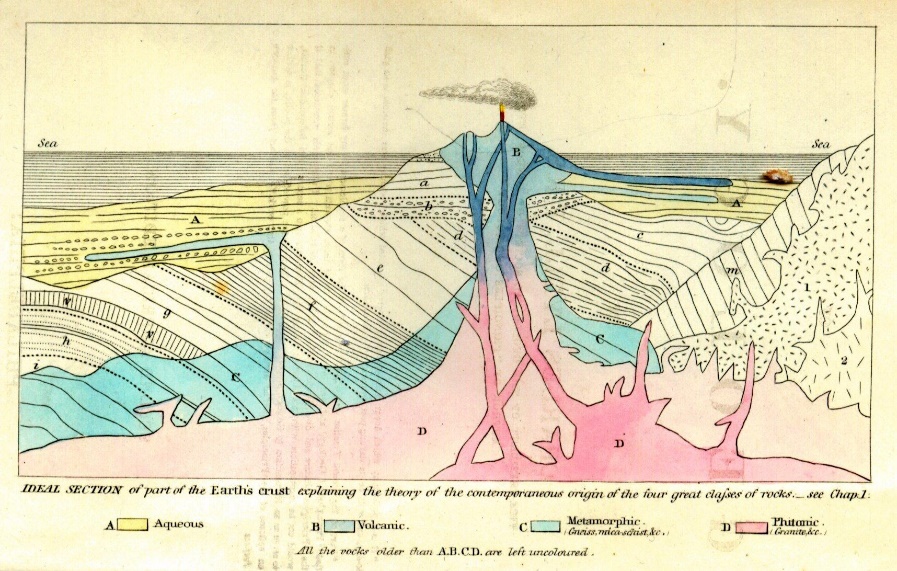
Another Scotsman, who was to become a highly influential geologist and a close friend of Darwin, was Charles Lyell (1797–1875). Lyell was originally a lawyer who began his studies of Geology at Oxford under the tutelage of catastrophist William Buckland, from whom he diverged when Buckland tried to find physical evidence of Noah’s flood from the Christian Bible. Lyell was instead intent on establishing geology as a science based on observation. Building upon Hutton’s ideas (published 50 years earlier), Lyell traveled throughout Europe, documenting evidence of uniformitarianism. During his travels, he cataloged evidence of sea level rise and fall and of volcanoes positioned atop much older rocks. He also found evidence of valleys formed through erosion, mountains resulting from earthquakes, and volcanic eruptions that had been witnessed or documented in the past (University of California Berkeley Museum of Paleontology n.d.). Lyell also espoused the principle that “rocks and strata (layers of rock) increase in age the further down they are in a geological sequence. Barring obvious upheavals or other evidence of disturbance, the same principle must apply to any fossils contained within the rock. The lower down in a sequence of rocks a fossil is, the older it is likely to be (Wood 2005, 12).”
Lyell published the first edition of his three-volume Principles of Geology in 1830–1833 (Figure 2.7). It established geology as a science, underwent constant revisions as new scientific evidence was discovered, and was published in 12 editions during Lyell’s lifetime. In it, he espoused the key concept of uniformitarianism—that “the present is the key to the past.” What this meant was that geological remains from the distant past can be explained by reference to geological processes now in operation and directly observable.
Jean-Baptiste Lamarck (1744–1829) was the first Western scientist to propose a mechanism explaining why and how traits changed in species over time, as well as to recognize the importance of the physical environment in acting on and shaping physical characteristics. Lamarck’s view of how and why species changed through time, known as the “Theory of Inheritance of Acquired Characteristics,” was first presented in the introductory lecture to students in his invertebrate zoology class at the Museum of Natural History in Paris in 1802 (Burkhardt 2013). It was based on the idea that as animals adapted to their environments through the use and disuse of characteristics, their adaptations were passed on to their offspring through reproduction (Figure 2.8). Lamarck was right about the environment having an influence on characteristics of species, as well as about variations being passed on through reproduction. He simply had the mechanism wrong.


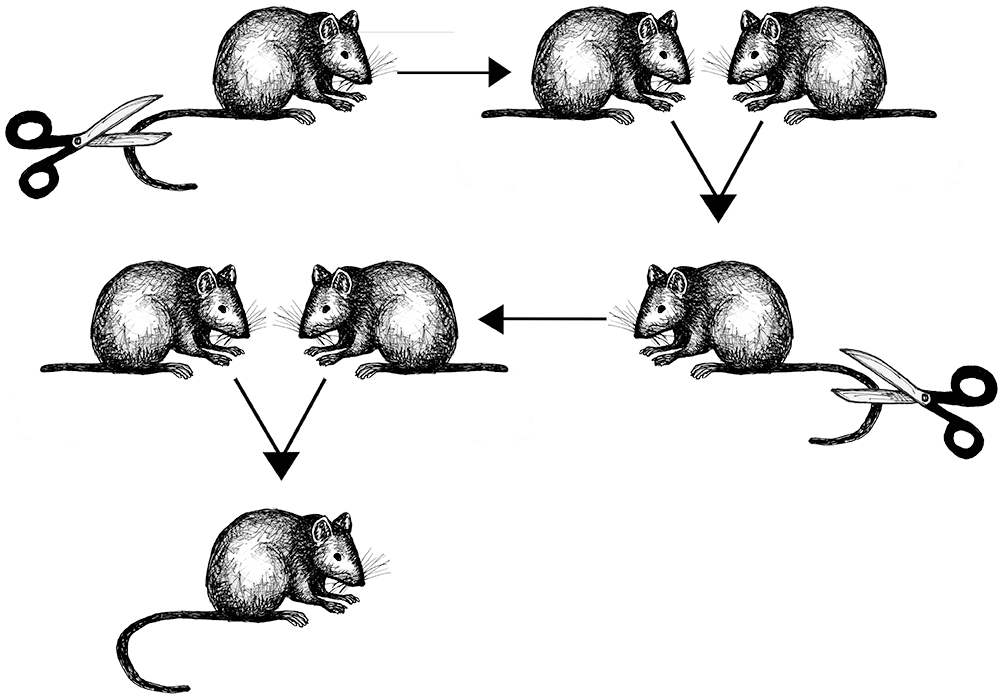
Lamarck’s theory involved a three-step process. Step one involves an animal experiencing a radical change in its environment. Step two is the animal (either individual or species) responding with a new kind of behavior. Step three is how the behavioral change results in morphological (meaning physical) changes to the animal that are successfully passed on to subsequent generations (Ward 2018, 8). Lamarck’s most famous example was the proposition that giraffes actively stretched their necks to reach leaves on tall trees to eat. Over their lifetimes, the continuation of this habit resulted in gradual lengthening of the neck. These longer necks were then passed on to their offspring. Lamarck’s theory was disproved when evolutionary biologist August Weismann published the results of an experiment involving mice (Figure 2.9). Weismann amputated the tails of 68 mice and then successively bred five generations of them, removing the tails of all offspring in each generation, eventually producing 901 mice, all of whom had perfectly healthy long tails in spite of having parents whose tails were missing (Weismann 1889).
How giraffes actually ended up with long necks is a different story. In an environment where the food supply is higher off the ground, and perhaps less available to competing species, giraffes who happened to have slightly longer necks (due to random individual variation and genetic mutation) would be more likely to survive. These giraffes would then be able to reproduce, passing along the slight variation in neck length that would allow their offspring to do the same. Over time, individuals with longer necks would be overrepresented in the population, and neck lengths overall would increase among giraffes. Unfortunately, Lamarck’s ideas challenged the scientific establishment of the time and were rejected. He was discredited and harassed “to the point of loss of money, reputation, and then health” (Ward 2018, 9).
The final piece in the evolutionary puzzle leading up to the theory of natural selection was put forth by Thomas Malthus (1766–1834), who published An Essay on Population in 1798. Malthus lived in England during the time of the Industrial Revolution. It was a time of great poverty and misery when many people migrated from the countryside to squalid, disease-ridden cities to work extremely long hours in dangerous conditions in factories, coal mines, and other industrial workplaces. Birth rates were high and starvation and disease were rampant. Malthus struggled to explain why. His answer was basically the idea of carrying capacity, an ecological concept still in use today. Malthus suggested the rate of population growth exceeded the rate of increase of the human food supply. In other words, people were outgrowing the available food crops. He also suggested that populations of animals and plants were naturally constrained by the food supply, resulting in reductions in population in times of scarcity, “restraining them within the prescribed bounds” (Moore 1993, 147). But, despite significant challenges, some individuals always survived. This was the key to later understandings of evolutionary change in species over time.
Exercises
The Journey to Natural Selection
In Western European thought, the individual most closely associated with evolution is Charles Darwin (1809–1882). However, as one can see from the individuals and ideas presented in the prior section, he was not the first person to explore evolution and how it might work. In fact, Darwin built upon and synthesized many of the ideas—from geology to biology, ecology, and economy—discussed above. He was simply in the right place at the right time. If he had not worked out his ideas when he did, someone else would have. As a matter of fact, as noted below, someone else did, forcing Darwin to publicly reveal his theory.
Darwin’s journey to the discovery of natural selection began during a childhood spent being curious, experimenting, and collecting natural specimens. When Darwin was 12 years old, his nickname was “Gas” because of the foul-smelling chemistry experiments he and his older brother, Erasmus, performed late into the evenings in their makeshift laboratory in the garden of their parent’s home (Costa 2017). Darwin was also a lifelong collector of biological and geological specimens, most famously beetles, at times going to great lengths in pursuit of a new specimen, as one of his personal letters relates,
I will give a proof of my zeal: one day, on tearing off some old bark, I saw two rare beetles, and seized one in each hand; then I saw a third and new kind, which I could not bear to lose, so that I popped the one which I held in my right hand into my mouth. Alas! it ejected some intensely acrid fluid, which burnt my tongue so that I was forced to spit the beetle out, which was lost, as was the third one. [Darwin 2001 (1897), 50].
Darwin continued his observations and experiments during his formal education, culminating in his graduation from Cambridge in 1831, at which point he was invited to become a gentleman naturalist for a British Royal Navy surveying mission of the globe aboard the H.M.S. Beagle. It is worth noting that Darwin was only 22 years old and the captain’s third choice for the position (Costa 2017), but he proved extremely curious and methodical. The mission departed in December of 1831 and returned five years later (Figure 2.10). During this time, Darwin produced copious notebooks, observations, drawings, and reflections on the natural phenomena he encountered and the experiments he performed.
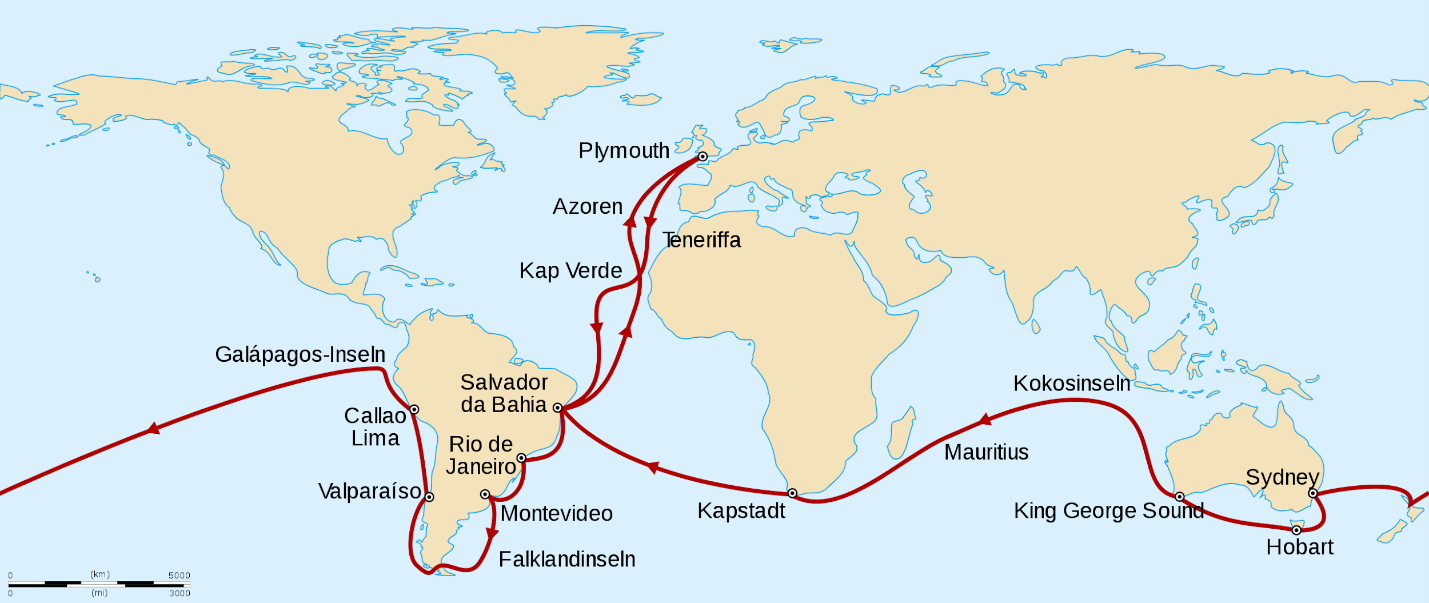
Discussing all of Darwin’s work aboard the Beagle is beyond the scope of this chapter, but his primary interests were in cataloging new varieties of plant and animal life and examining the geology of the places the ship made landfall. Part of Darwin’s success with regard to both ventures was due to his extreme seasickness, which began before the ship even left Plymouth Harbor. It never let up, encouraging Darwin to go ashore at every available opportunity. “In fact, of the nearly five years of the voyage, Darwin was actually on board the ship for just a year and a half altogether” (Costa 2017, 18).
During the voyage, the young Darwin tried to make sense of what he saw through the lens of the scientific paradigms he held when he left England, but he continually made observations that challenged these paradigms. For example, while the Beagle crewmen were charting the coast of Argentina, Darwin conducted fieldwork on land. There he observed species that were new to him, like armadillos. He also collected fossils, including those of extinct armadillos. Meaning, he had found both extant and extinct members of the same species in the same geographic location, which challenged the theory of catastrophism put forth by Cuvier, who argued that each variant of an animal, living or extinct, was its own distinct species (Moore 1993, 144). Darwin also observed geographic variation in the same species all along the east coast of South America, from Brazil to the southern tip of Argentina. He noted that some species were found in multiple localities and differed from place to place. Those living closer to each other exhibited only slight variations, while those living further apart might be cataloged as entirely different species if one did not know better.
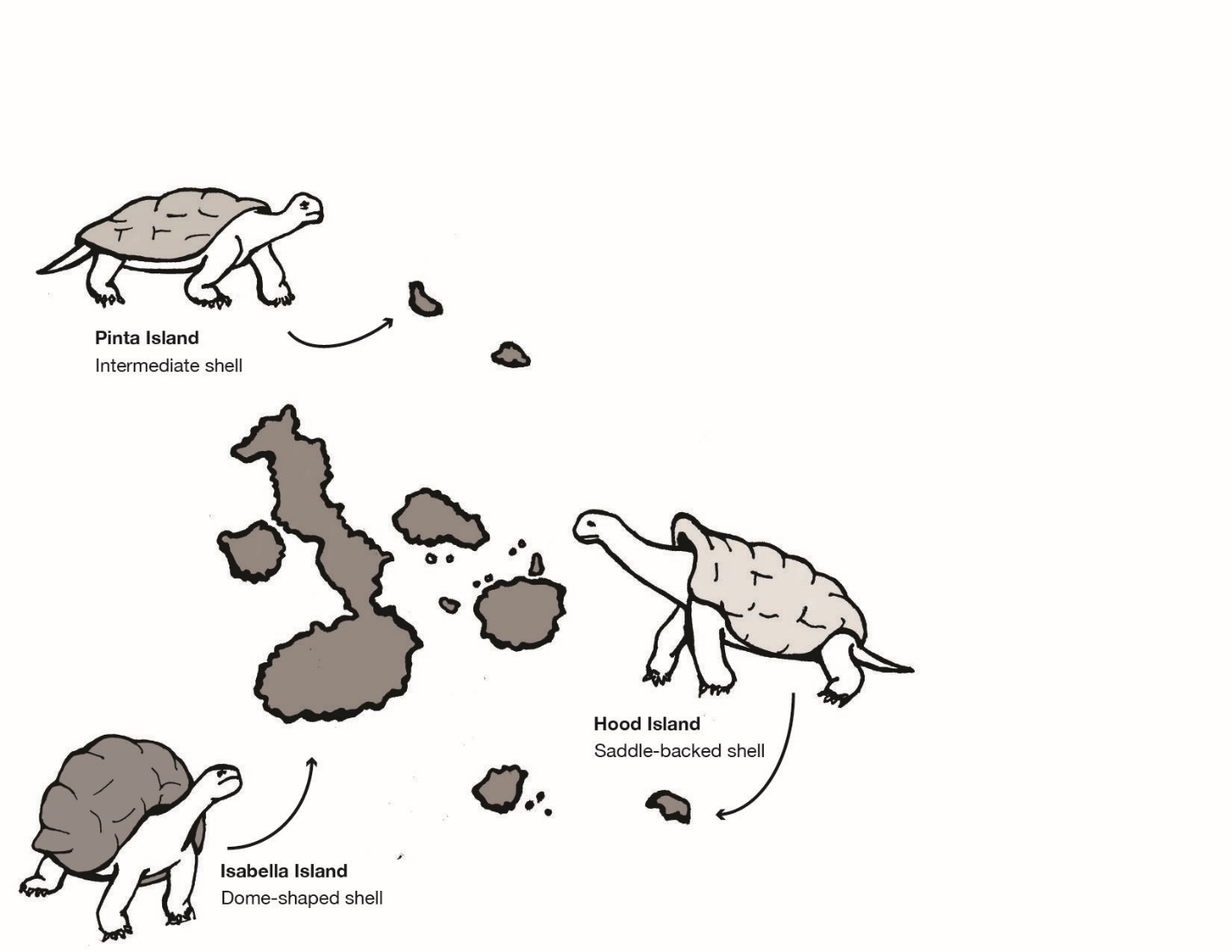
He made similar observations in the Galapagos Islands located off the northwest coast of Ecuador, with regard to giant tortoises and finches (Figure 2.11). A local resident of the islands explained to Darwin that each island had its own variety of tortoise and that locals could discern which island a tortoise came from simply by looking at it. Darwin noted other such examples in both plants and animals, meaning geographic variation was occurring on separate, neighboring islands.
Prevailing views of time argued that variations in living species, and even the fossil armadillos and the living armadillos, were the result of separate creation events. According to this view, each variation, no matter how slight, was a different species. Challenging these ideas would mean challenging not only catastrophism, but the Fixity of Species and other well-accepted ideas of the time. Darwin was aware that he was a young, unestablished naturalist. He was also aware of the ruin that befell Lamarck when his theories were rejected. Lastly, Lyell, who was a good friend of Darwin’s, rejected evolution altogether. It is no wonder that Darwin published a great deal about the geological and fossil data he collected when he returned from the voyage, but not his early hypotheses about evolution.
Exercises
Upon Darwin’s return to England, it took another twenty years of data collection and experimentation before he was ready to share his conclusions about evolution. Much of this work was conducted at Down House, his home of forty years, where he performed all sorts of experiments that laid the groundwork for his ideas about evolution. Darwin’s home was his laboratory, and he engaged the help of his children, neighbors, friends, and servants in collecting, dissecting, and experimenting. At one point in the 1850s, sheets of moistened paper covered with frogs eggs lined the hallways of the house, while flocks of sixteen different pigeon breeds cooed outside, glass jars filled with salt water and floating seeds filled the cellar, and the smell of dissected pigeon skeletons pervaded the air inside the house. There were also ongoing experiments in the yard, including piles of dissected flowers, beekeeping, and fenced-off plots of land where seedlings were under study. Darwin was a keen experimental scientist, observer, and a prolific writer and presenter of scientific papers. He regarded his work as “one long argument” that never really ended. In fact, Darwin published ten books after On the Origin of Species, addressing such far-ranging topics as animal behavior, orchids, and domestication, among others (Costa 2017).
Darwin may not have published Origins in 1859 had it not been for receiving a paper in June of 1858 from Alfred Russel Wallace, an English naturalist working in Malaysia, espousing the same ideas. Wallace had sent the paper to Darwin asking if it was worthy of publication and requesting he forward it to Lyell and the English botanist, Joseph Hooker. Darwin wrote to Lyell and Hooker about Wallace’s paper, entitled On the Tendency of Varieties to Depart Indefinitely from the Original Type. In recognition that both Wallace and Darwin had arrived at the same discovery, a “joint” paper composed of four parts (none of them actually coauthored) was read to the Linnaean Society by Lyell, then secretary of the Society, on July 1, 1858, and published on August 20. Darwin published On the Origin of Species 15 months later. (The original composite paper read before the Linnaean society is available to read for free from the Alfred Russell Wallace Website, on the 1858 Darwin-Wallace paper page.)
Exercises
The Mechanism of Natural Selection
On the subject of natural selection and how it works, let’s hear from Darwin himself from the original publication of On the Origin of Species (1859):
A struggle for existence inevitably follows from the high rate at which all organic beings tend to increase. Every being, which during its natural lifetime produces several eggs or seeds, must suffer destruction during some period of its life, and during some season or occasional year, otherwise, on the principle of geometric increase, its numbers would quickly become so inordinately great that no country could support the product. Hence, as more individuals are produced than can possibly survive, there must in every case be a struggle for existence…It is the doctrine of Malthus applied with manifold force to the whole animal and vegetable kingdoms…There is no exception to the rule that every organic being naturally increases at so high a rate, that if not destroyed, the earth would soon be covered by the progeny of a single pair. …Owing to this struggle for life, any variation, however slight and from whatever cause proceeding, if it be in any degree profitable to an individual of any species…will tend to the preservation of that individual, and will be inherited by its offspring. The offspring, also, will thus have a better chance of surviving, for, of the many individuals of any species which are periodically born, but a small number can survive. I have called this principle, by which slight variation, if useful, is preserved, by the term Natural Selection. [Darwin 1859, 61–62]
Let us take a moment here to explore the mechanism of natural selection in more detail. Before we begin, it is important to recognize that Darwin defined evolution as descent with modification, by which he meant that species share a common ancestor yet change over time, giving rise to new species. Descent with modification refers to the fact that offspring from two parents look different from each of their parents, and from each other, meaning they descend with slight differences (“modifications”). If you have ever observed a litter of puppies or a field of flowers and stopped to examine each individual closely, you have seen that each differs from the next, and none look exactly like their parents. These variations are random, not specific, and may or may not be present in the following generations.
Darwin struggled to explain why some slight differences were preserved over time, while others were not. He turned to what he knew of animal breeding (artificial selection) for an explanation (Richards 1998). Darwin bred different breeds of pigeons at Down House, carefully documenting phenotypic differences across generations, including slight anatomical variations he observed through dissection. He also grew and crossbred species of flowers and dissected those too. Darwin was also very fond of hunting and of hunting dogs. In an early draft of his theory on speciation, he used greyhounds as an example of adaptation and selection, “noting how its every bone and muscle, instinct and habit, were fitted to run down hare (rabbits) (University of Cambridge n.d.).” In each case of plant and animal breeding Darwin observed, he noted that humans were selecting variants in each generation that had characteristics humans desired (i.e., sweetness of fruits, colors of flowers, fur type and color of animals). Breeders then continually bred plants and animals with the desired variants, over and over again. These small changes added up over time to create new species of plants and breeds of animals. Darwin also noted that artificial selection does not necessarily render plants or animals better adapted to their original environments. The characteristics humans desire often result in plants less likely to survive in the wild and animals more likely to suffer from certain behavioral or health problems. One has only to examine high rates of hip dysplasia in several modern breeds of dogs to observe what Darwin was referring to.
From his studies of artificial selection, Darwin drew the conclusion that nature also acts upon variations among members of the same species. Instead of human intervention, the forces of nature, such as heat, cold, predation, disease, and now climate change, determine which offspring, with which variants, survive and reproduce. These individuals then pass down these favorable variants to their own offspring. In this way, nature selects for traits that are beneficial within a particular environment and selects against traits that are disadvantageous within a particular environment. Over many generations, populations of a species become more and more adapted (or, in evolutionary terms, “fit”) for their specific environments. Darwin named this process natural selection.
This theory explained the variations in tortoises Darwin had observed years earlier in the Galapagos Islands (see Figure 2.11). Tortoises who lived on larger islands with lush vegetation to feed on were larger than those on smaller islands. They also had shorter necks and dome-shaped shells as their food was close to the ground. Tortoises on smaller, drier islands fed on cacti, which grew much taller. These tortoises had longer necks, longer front legs, and saddle-shaped shells, which allowed them to successfully stretch to reach the edible cactus pads that grew on the tops of the plants. How did these observable differences in the two tortoise populations emerge? Darwin would argue that, over time, small, random variations in the tortoises were differentially selected for by the distinct natural environments on different islands.
In addition to the biogeographical evidence Darwin offered from his research aboard the Beagle, as well as the evidence he documented from the artificial selection of plants and animals, he also relied, where possible, on fossil evidence. One example, mentioned above, were the fossil findings of extinct armadillos in Argentina in the same locations as living armadillos. Unfortunately, as Darwin himself noted, the geological record was incomplete, most often missing the transitional fossil forms that bridge extinct and living species. That issue has since been resolved with scientific research in geochronology and paleontology, among other fields. It is now well-established that life is far more ancient than was believed in Darwin’s time and that these ancient forms of life were the ancestors to all life on this planet (Kutschera and Niklas 2004).
What Darwin Was Missing
Although the theory of evolution by natural selection gained traction in scientific circles in the decades after Darwin’s publication of Origins, he was never able to discover the mechanisms that caused variation within members of the same species or the means by which traits were inherited. This began later in 1892 with the publication of The Germ-Plasm: A Theory of Heredity by August Weismann, the same Weismann of the mouse tail experiment presented earlier in this chapter. In his book, Weissman proposed a theory of germ-plasm, which was a precursor to the later discovery and understanding of DNA. Weismann specialized in cytology, a branch of biology devoted to understanding the function of plant and animal cells. Germ-plasm, he proposed, was a substance in the germ cells (what we would call gametes, or sex cells, today) that carried hereditary information. He predicted that an offspring inherits half of its germ-plasm from each of its parents, and claimed that other cells (e.g. somatic, or body, cells) could not transmit genetic information from parents to offspring. This thereby erased the possibility that acquired traits (which he argued resided in somatic cells) could be inherited (Zou 2015). This contribution to evolutionary theory was an important step toward understanding genetic inheritance, but a connection between genetics and evolution was still lacking.
A series of lectures by a deceased Augustinian monk named Gregor Mendel (1822–1884), originally published in 1865, changed that perspective (Moore 1993, 285). Although Darwin was unknown to Mendel, he began a series of experiments with pea plants shortly after the publication of Darwin’s Origins, aiming to add to evolutionary understandings of heredity. As Mendel bred different generations of pea plants that had differences in seed shape and color, pod shape and color, flower position, and stem length, he documented consistent expression of some variations over others in subsequent generations. He meticulously documented the statistics of each crossing of plants and the percentages of phenotypes that resulted, eventually discovering the concept of dominance and recessiveness of characteristics. He also documented that there is no blending of inherited characteristics. For example, pea pod colors in the offspring of two parent plants, one with yellow pods and one with green, were either yellow or green, not yellowish green. Mendel also discovered that characteristics are inherited and expressed independently of each other, meaning the color of the pea pod was not necessarily expressed in conjunction with the pod being wrinkled or smooth. The recognition of the importance of Mendel’s work began with its rediscovery by Hugo de Vries and Carl Correns, both of whom were working on hypotheses regarding heredity in plants and had arrived at conclusions similar to Mendel’s. Both published papers supporting Mendel’s conclusions in 1900 (Moore 1993). Research into the inheritance of characteristics continued through the next three decades, and by the close of the 1930s, no major scientific questions remained regarding the transmission of heredity through genes, although what genes did and what chemicals they were made of were still under investigation.
Exercises
The Modern Synthesis refers to the merging of Mendelian genetics with Darwinian evolution that took place between 1930 and 1950. The basic principles of the synthetic theory were influenced by scientists working in many different fields, including genetics, zoology, biology, paleontology, botany, and statistics. Although there were differences of opinion among them, evolution came to be defined as changes in allele frequencies within populations. Genetic mutations, changes in the genetic code that are the original source of variation in every living thing, were believed to be random, the sources of phenotypic variation, and transmitted through sexual reproduction. These assertions were supported by a growing body of field and laboratory research, as well as new work in mathematics in the field of population genetics that defined evolution as numerical changes in gene frequencies within an interbreeding population from one generation to the next (Corning 2020). These changes in gene frequencies were argued to be the result of natural selection, mutation, migration (gene flow), and genetic drift, or random chance. Empirical research and mathematics demonstrated that very small selective forces acting over a relatively long time were able to generate substantial evolutionary change, including speciation (Plutynski 2009). Thus, the Modern Synthesis encompassed both microevolution, which refers to changes in gene frequencies between generations within a population, and macroevolution, longer-term changes in a population that can eventually result in speciation, wherein individuals from different populations are no longer able to successfully interbreed and produce viable offspring.
Exercises

Genetics and the Origin of Species, published in 1937 by Theodosius Dobzhansky (Figure 2.12), was a cornerstone of the modern synthesis, applying genetics to the study of natural selection in wild populations, appealing to both geneticists and field biologists. Dobzhansky was interested in speciation, particularly in finding out what kept one species distinct from another and how speciation occurred. His research involved fruit flies, the species Drosophila pseudoobscura. At the time he began in the 1920s, biologists assumed all members of the same species had nearly identical genes. Dobzhansky traveled from Canada to Mexico capturing wild members of D.pseudoobscura, discovering that different populations had different combinations of alleles (forms of a gene) that distinguished them from other populations, even though they were all members of the same species. What, then, led to the creation of new species? Dobzhansky realized it was sexual selection. Members of the same species are more likely to live among their own kind and to recognize, and prefer, them as mates. Over time, as a result of random mutations, natural selection in a given environment, and genetic drift, meaning random changes in allele frequencies, members of the same population accumulate mutations distinct to their own gene pool, eventually becoming genetically distinct from other populations. What this means is that they have become a new species.
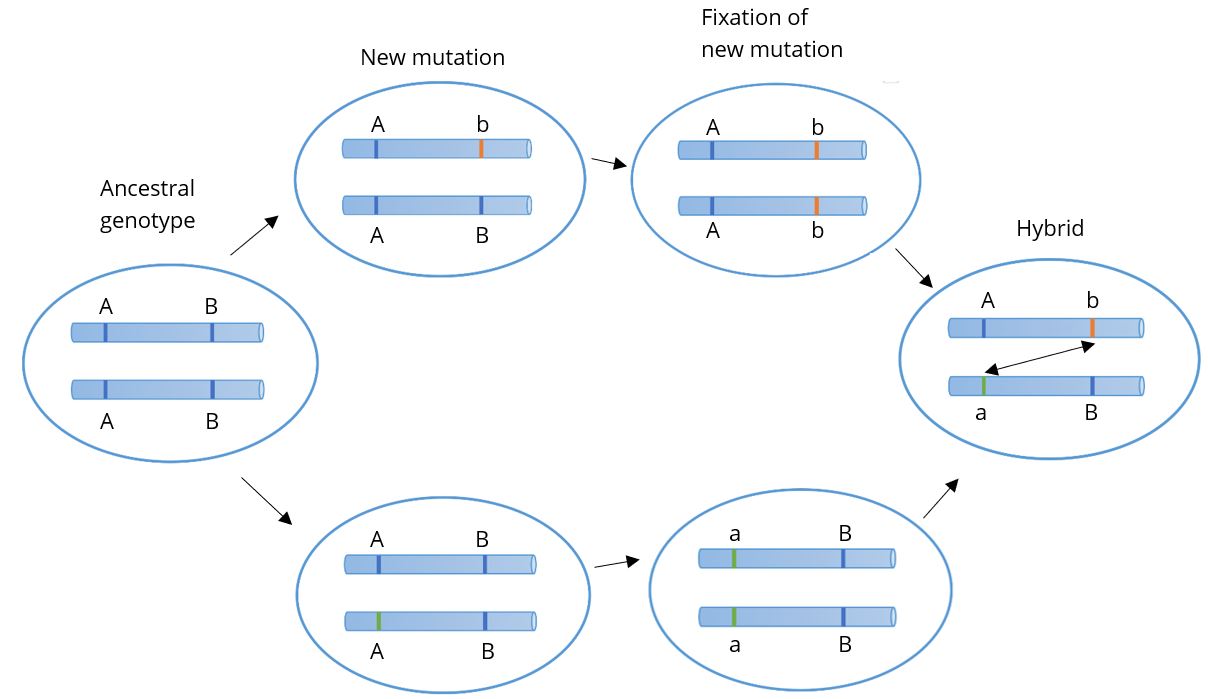
From these studies, Dobzhansky and others developed the Bateson-Dobzhansky-Muller model, also known as Dobzhansky-Muller model (Figure 2.13). It is a model of the evolution of genetic incompatibility. Combining genetics with natural selection, the model is important in understanding the role of reproductive isolation during speciation and the role of natural selection in bringing it about. Due to sexual selection (mate preference), populations can become reproductively isolated from one another. Eventually, novel mutations may arise and be selected for in one or both populations, rendering members of each genetically incompatible with the other, resulting in two distinct species.
Are We Still Evolving?
Change in species or evolution is always studied in terms of populations and not individuals. If a population were to stay exactly the same from one generation to the next, it would not be evolving. If allele frequencies change, then, yes, that population is evolving. Alleles are just variants of genes (more on this later). So all that needs to change is the number or ratio of gene variants. We don’t have to develop a third eye or spidey senses to be evolving. So we can ask, do frequencies of alleles change in human populations? Of course they do. Absolutely.
We sometimes see organs or structures that once had a function or have changed somewhat in function or some structures that are disappearing altogether. Sometimes, these are called vestigial structures. Wisdom teeth are already changing. Joylin Namie in Chapter 2 “History of Evolutionary Thought” in Explorations, An Open Invitation to Biological Anthropology explains. Wisdom teeth are also undergoing evolutionary pressure. Have you or anyone in your family had their wisdom teeth removed? While it can be a painful and expensive process, it is a common experience in Western nations. It begs the question as to why there is no longer room in our mouths for all of our teeth? Biological anthropologist Daniel Lieberman offers several reasons, including that modern humans are growing faster and maturing earlier, which could be leading to impaction if skeletal growth takes place faster than dental growth. He also argues that the soft diets many modern humans consume generate insufficient strain to stimulate enough growth in our jaws to accommodate all of our teeth. Lastly, as the human brain has expanded over the past hundreds of thousands of years, it is taking up more space in the skull, causing the jaw to shrink, leaving no room for third molars (Lieberman 2011).
Conversely, do you know anyone whose wisdom teeth never came in? That is fairly common in some populations, suggesting evolutionary pressure favoring the absence of wisdom teeth has been culturally influenced. The oldest fossil evidence of skulls missing third molars was found in China and is 300,000 to 400,000 years old, suggesting the earliest mutation selecting against the eruption of wisdom teeth arose in Asia (Main 2013). Since that time, jaws have continued to decrease in size to the point they often cannot accommodate third molars, which can become impacted, painful, and even infected, a condition physical anthropologist Alan Main argues might have interfered with the ability to survive and reproduce in ancestral populations (Main 2013). As we have learned, a mutation that positively influences reproductive success—such as being born without the trait to develop wisdom teeth—would likely be selected for over time. Evidence in modern humans suggests that this is the case, with 40% of modern Asians and 45% of Native Alaskans and Greenlanders (populations descended from Asian populations) lacking wisdom teeth. The percentage among those of European descent ranges from 10 to 25% and for African Americans is 11% (Main 2013).
Key Terms
Allele: A nonidentical DNA sequence found in the same gene location on a homologous chromosome, or gene copy, that codes for the same trait but produces a different phenotype.
Artificial selection: The identification by humans of desirable traits in plants and animals, and the subsequent steps taken to enhance and perpetuate those traits in future generations.
Binomial nomenclature: A system of classification in which a species of animal or plant receives a name consisting of two terms: the first identifies the genus to which it belongs, and the second identifies the species.
Carrying capacity: The number of living organisms, including animals, crops, and humans, that a geographic area can support without environmental degradation.
Catastrophism: The theory that the Earth’s geology has largely been shaped by sudden, short-lived, violent events, possibly worldwide in scope. Compare to uniformitarianism.
Comparative anatomy: Georges-Louis Leclerc’s technique of comparing similar anatomical structures across different species.
Creationism: The belief that the universe and all living organisms originate from specific acts of divine creation, as in the Biblical account, rather than by natural processes such as evolution.
Empiricism: The idea that all learning and knowledge derives from experience and observation. It became prominent in the 17th and 18th centuries in western Europe due to the rise of experimental science.
Evolution: In a biological sense, this term refers to cumulative inherited change in a population of organisms through time. More specifically, evolution is defined as a change in allele (gene) frequencies from one generation to the next among members of an interbreeding population.
Extant: Still in existence; surviving.
Extinct: Said of a species, family, or other group of animals or plants that has no living members; no longer in existence.
Fixity of Species: The idea that a species, once created, remains unchanged over time.
Gene: A sequence of DNA that provides coding information for the construction of proteins.
Genetic drift: Random changes in allele frequencies within a population from one generation to the next.
Gene flow: The introduction of new genetic material into a population through interbreeding between two distinct populations.
Gene pool: The entire collection of genetic material in a breeding community that can be passed from one generation to the next.
Genotype: The genotype of an organism is its complete set of genetic material—its unique sequence of DNA. Genotype also refers to the alleles or variants an individual carries in a particular gene or genetic location.
Hybrid: Offspring of parents that differ in genetically determined traits.
Intelligent design: A pseudoscientific set of beliefs based on the notion that life on earth is so complex that it cannot be explained by the scientific theory of evolution and therefore must have been designed by a supernatural entity.
Macroevolution: Large and often-complex changes in biological populations, such as species formation.
Microevolution: Changes in the frequency of a gene or allele in an interbreeding population.
Modern synthesis: The mid–20th century merging of Mendelian genetics with Darwinian evolution that resulted in a unified theory of evolution.
Natural selection: The natural process by which the survival and reproductive success of individuals or groups within an interbreeding population that are best adjusted to their environment leads to the perpetuation of genetic qualities best suited to that particular environment at that point in time.
Phenotype: The detectable or visible expression of an organism’s genotype.
Scientific method: A method of procedure that has characterized natural science since the 17th century, consisting of systematic observation, measurement, experimentation, and the formulation, testing, and modification of hypotheses.
Speciation: The process by which new genetically distinct species evolve from the main population, usually through geographic isolation or other barriers to gene flow.
Species: A group of living organisms consisting of similar individuals capable of exchanging genes or interbreeding. The species is the principal natural taxonomic unit, ranking below a genus and denoted by a Latin binomial (e.g., Homo sapiens).
Uniformitarianism: The theory that changes in the earth’s crust during geologic history have resulted from the action of continuous and uniform processes—such as wind, precipitation, evaporation, condensation, erosion, and volcanic action—that continue to act in the present. Compare to catastrophism.
things in life are teaching, traveling, and spending time with her dog, Charley.
For Further Exploration
Costa, James T. 2017. Darwin’s Backyard: How Small Experiments Led to a Big Theory. New York: W.W. Norton.
Darwin, Charles. 1905. The Voyage of the Beagle. (Originally published in 1839 as Journal and Remarks). [Author’s note: Several editions exist with different publishers, including illustrated editions, paperback editions, and e-books.]
Moore, John A. 1993. Science as a Way of Knowing: The Foundations of Modern Biology. Cambridge, MA: Harvard University Press.
Image Descriptions
Figure 2.10: A simple world map with a line depicting the ocean route that the H.M.S. Beagle took, and the ports visited. Leaving Plymouth (Europe), the ship traveled south past Africa, around most of the coastline of South America, then around the southern sides of Australia and Africa, before returning briefly to South America and then Europe. Ports noted on the map include Plymouth (Europe); Teneriffa (Africa), Kap Verde, Salvador da Bahia, Rio de Janeiro, Montevideo, Falklandinseln, Valparaiso, Callao Lima, Galápagos-Inseln Kap Verde (South America); Sydney, Hobart, King George Sound (Australia), Kokosinseln, Mauritius (Southeast Asian Islands); Kapstadt (Africa); Salvador da Bahia (South America); Azoren (Atlantic Island); Plymouth (Europe).
Attribution
This chapter is from the following source with section deletions, minor additions, and minor editing.
Joylin Namie, “A History of Evolutionary Thought,” Explorations: An Open Invitation to Biological Anthropology, 2nd Edition edited by Beth Shook, Lara Braff, Katie Nelson, Kelsie Aguilera, licensed under a Creative Commons Attribution-NonCommercial 4.0 International License.
In a biological sense, this term refers to cumulative inherited change in a population of organisms through time. More specifically, evolution is defined as a change in allele (gene) frequencies from one generation to the next among members of an interbreeding population.
The natural process by which the survival and reproductive success of individuals or groups within an interbreeding population that are best adjusted to their environment leads to the perpetuation of genetic qualities best suited to that particular environment at that point in time.
A method of procedure that has characterized natural science since the 17th century, consisting of systematic observation, measurement, experimentation, and the formulation, testing, and modification of hypotheses.
The idea that all learning and knowledge derives from experience and observation. It became prominent in the 17th and 18th centuries in western Europe due to the rise of experimental science.
A system of classification in which a species of animal or plant receives a name consisting of two terms: the first identifies the genus to which it belongs, and the second identifies the species.
Said of a species, family, or other group of animals or plants that has no living members; no longer in existence.
The theory that the Earth’s geology has largely been shaped by sudden, short-lived, violent events, possibly worldwide in scope. Compare to uniformitarianism.
The number of living organisms, including animals, crops, and humans, that a geographic area can support without environmental degradation.
Still in existence; surviving.
The idea that a species, once created, remains unchanged over time.
The identification by humans of desirable traits in plants and animals, and the subsequent steps taken to enhance and perpetuate those traits in future generations.
The mid–20th century merging of Mendelian genetics with Darwinian evolution that resulted in a unified theory of evolution.
The introduction of new genetic material into a population through interbreeding between two distinct populations.
Random changes in allele frequencies within a population from one generation to the next.
Changes in the frequency of a gene or allele in an interbreeding population.
Large and often-complex changes in biological populations, such as species formation.
The process by which new genetically distinct species evolve from the main population, usually through geographic isolation or other barriers to gene flow.
Variant forms of genes.
A sequence of DNA that provides coding information for the construction of proteins.
The entire collection of genetic material in a breeding community that can be passed from one generation to the next.
A group of living organisms consisting of similar individuals capable of exchanging genes or interbreeding. The species is the principal natural taxonomic unit, ranking below a genus and denoted by a Latin binomial (e.g., Homo sapiens).