
5 Paleoanthropology
Paleoanthropology
Paleoanthropology is a specialized field in biological anthropology that is interested in human origins. This subject is investigated through two main avenues—the study of ancient fossils and genetics. Paleoanthropologists group ancient species into taxonomic groups (just like modern primates are classified in taxonomic groups) according to their morphology and sometimes their genetic makeup. This is done in order to understand the evolution, or change over time, in the human species.
Fossils are simply the remains of once-living organisms, whose tissues (usually bones and teeth) have been replaced by rock. Fossils only form when the organism was quickly buried and conditions are ideal. As a result, the fossil record represents only a small fraction of the populations that were once living. Paleoanthropologists also group species based on their genetics. This approach is only possible when tissues have not been fully mineralized or fossilized. This sparsity of the fossil record and the lack of preserved genetic material makes paleoanthropology a challenging discipline.
The processes by which species diversify today, as seen in the rock pocket mouse example, were also at play in the past. Mutations occur randomly and processes like natural selection and founder effect lead to diversification in species. These processes can lead to many branches that derive from a common ancestor. Change over time does not happen in a linear step-wise fashion, but rather in a branching fashion. In this chapter, we will look at some of those species that are thought to be ancestral to humans and why they are considered to be in the human family.
The Problem of Time
It was once thought that there wasn’t much time for a lot to happen on planet earth. Many scholars attempted to calculate the age of the Earth and/or people on the Earth using the available texts. Archbishop James Ussher’s calculation used, in part, the genealogies and reigns outlined in the Bible along with known dates of rulers. Ussher calculated the beginning of creation to 6004 BP. These calculations were taken very seriously, and even Sir Isaac Newton attempted an estimate.
While Usher’s estimate was extremely influential, the physical evidence did not align well with this relatively short time frame. In the 1790s, John Frere, great-great-grandfather of famous paleoanthropologist Mary Leakey, excavated stone tools four meters (that’s about 12 feet) below the ground surface in ancient lake deposits in Suffolk, England. How did these artifacts get to be so deep in the ground, he wondered? Going against conventional thought, Frere concluded, “The situation in which these weapons were found may tempt us to refer them to a very ancient period indeed.” A similar situation was coming to light in France. In the 1840s, a customs official named Boucher de Perthes uncovered stone tools associated with extinct elephant remains (mammoth) deep in ancient Somme River gravels in France. Like Frere, he concluded that the human presence in Europe must be much older than previously thought.
The problem was that people were finding artifacts and fossils in places where conventional thinking said they shouldn’t be. The deeper the geological layers, the older the fossils. These layers and the fossils they contain provide a blueprint for how life changed on earth since its beginnings around 4.5 billion years ago. If the earth’s history were scaled to a year, humans appear in the last hour of the year. We now know, based on geology and paleontology, that the earth is much older than previously believed and that humans and our cousins are relatively late arrivals.
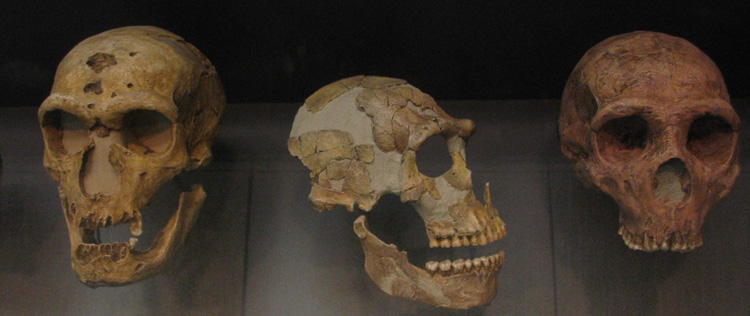
So what kinds of things do paleoanthropologists look for in the fossil record? Paleoanthropologists of course, look for fossils that have features of primates. In addition, they look for morphological traits and behaviors that stand out as human compared to other primates. Bipedalism (walking on two legs), large brain to body size, a reduced face, and changes in dentition (teeth) are features that are unique to humans in the primate order, and so paleoanthropologists use these to identify human ancestors, or hominins. All hominins are bipedal, and all are primates, so we can think of them as bipedal primates. Hominins include our extinct bipedal ancestors and relatives since our divergence from the last common ancestor (LCA) that we share with chimpanzees. Other human-like features like large brains, a reduced face, and changes in dentition appear later in the fossil record. That is, our earliest ancestors walked upright but had ape-like teeth and relatively small brains.
Bipedalism is an important concept in the study of hominins. Chimps, bonobos, and gorillas can walk on two legs, but they are facultative bipeds, meaning they can walk on two legs for short periods. More typically, they propel themselves forward with their back legs and support the front part of their bodies with their arms on curled-up hands—knuckle-walking. Humans, unlike apes, are obligate bipeds, meaning we can walk bipedally for long periods or habitually. Bipedalism is so quintessentially human that you can put two legs on a sponge (Spongebob) or a fish (Darwin on Gumball), and they become recognizably human-like.

An early discovery called Piltdown Man reveals early assumptions about which human-like traits must have come first. Many assumed that large brains must have originated first, and then our species would be motivated to walk on two legs. Piltdown Man was discovered in 1912 in England by Charles Dawson who claimed he had found the proverbial “missing link” between apes and humans. Piltdown Man had a large brain but an ape-like jaw, exactly what some scholars expected to find. It also was a feather in the cap for England for having the earliest example of a hominin. However, it quickly came to light that Piltdown Man was actually an ancient skull and a modern orangutan jaw made to look older. The teeth were also worn down to more closely resemble a human.

Even when fossil remains are not outright fraud, it is difficult to ascertain how closely related hominins are to each and to us. As discussed, taxonomy was based purely on form or morphology. Today, we also use genetics to under phylogeny or relatedness among primates. As is pointed out in the “Early Hominins” chapter in Explorations: An Open Invitation to Biological Anthropology, this is complicated when applied to living primates but becomes much more difficult when we try to categorize ancestor- descendant relationships for long-extinct species whose molecular information is no longer preserved. `We therefore find ourselves falling back on morphological comparisons, often of teeth and partially fossilized skeletal material. Cladistics groups organisms according to their last common ancestors based on shared derived traits. Remember, those are those that have been altered more recently. In the case of early hominins, these are often morphological traits that differ from those seen in earlier populations.
The authors of “Early Hominins” in Explorations: An Open Invitation to Biological Anthropology, explain different approaches to classifying hominin species. Species designations for early hominin specimens are often highly contested. This is due to the fragmentary nature of the fossil record, the large timescale (millions of years) with which paleoanthropologists need to work, and the difficulty in evaluating whether morphological differences and similarities are due to meaningful phylogenetic or biological differences or subtle differences/variation in niche occupation or time. In other words, do morphological differences indicate different species? How would classifying species in the paleoanthropological record compare with classifying living species today, for whom we can sequence genomes and observe lifestyles?
There are also broader philosophical differences among researchers when it comes to paleo-species designations. Some scientists, known as “lumpers,” argue that large variability is expected among multiple populations in a given species over time. These researchers will therefore, prefer to “lump” specimens of subtle differences into single taxa. Others, known as “splitters,” argue that species variability can be measured and that even subtle differences can imply differences in niche occupation that are extreme enough to mirror modern species differences. In general, splitters would consider geographic differences among populations as meaning that a species is polytypic (i.e., capable of interacting and breeding biologically but having morphological population differences). This is worth keeping in mind when learning about why species designations may be contested.
Bipedalism
All hominins are bipedal. Walking on two legs is the key defining feature of hominins, in addition of course to their other primate features. But how can paleoanthropologists determine if a species was bipedal or not? Several skeletal clues point to bipedality. First, the foramen magnum, literally “big hole,” from which the spinal cord exits at the base of the skull, is placed more anteriorly (more toward the front of the skull). This makes intuitive sense because the head sits atop the spine in bipeds.

Another indication of bipedality is something called the valgus knee, which is the angle of the knee between the femur and tibia. Having this inward angle allows for weight to be distributed around the center of gravity. This configuration allows us to shift our weight back and forth without leaning precipitously to one side. Latin dancing, like salsa and bachata, owes much to the valgus knee. The angling comes at some cost, though, because the knee is relatively weak, causing many people to experience knee pain. The valgus knee develops over time as children begin to walk. Chimps, being knucklewalkers, do not have a valgus knee, and no angling in.
Another indication of bipedality is a rather wide bowl-shaped pelvis in which the guts sit. The femur neck is angled to accommodate this configuration. Like the valgus knee, the rather long femoral neck is a point of weakness, leading to hip replacements in older individuals (Warren et al. “Early Hominins” chapter in Explorations: An Open Invitation to Biological Anthropology). You can see how elongated the chimpanzee pelvis is compared to the more bowl-like pelvis of hominins (Figure 5.5).

Bipeds also have an s-shaped spine, which helps to distribute body weight over the hips. Non-bipeds, like chimpanzees, do not need this Jenga-like balancing act. This stacking reduces the muscle engagement needed to keep us upight while walking, and therefore requires less engergy ( “Early Hominins” Explorations: An Open Invitation to Biological Anthropology, first edition). Like the valgus knee and long femoral neck, the s-shaped spine is often the source of lumbar pain and pinched nerves, as well as a source of income for chiropractors.
The biped foot is also much different than a quadruped or knucklewalker. Biped feet tend to be robust and large, having to carry the weight of the body. The heel is large and acts as a shock absorber. The big toe, or hallux, is also large in order to be used for “push off” while walking. In addition, biped feet tend not to be opposable or grasping, an adaptation to arboreality, but instead are rather rigid. Table 5.1 summarizes the signatures of bipedality in obligate bipeds.
Exercises
The authors of “Early Hominins” chapter in Explorations: An Open Invitation to Biological Anthropology, explain the emergence of bipedalism in early hominin species. Despite relatively few postcranial fragments, the fossil record in early hominins indicates a complex pattern of emergence of bipedalism. Key features, such as a more anteriorly placed foramen magnum, are argued to be seen even in the earliest discovered hominins, indicating an upright posture (Dart 1925). Some early species appear to have a mix of ancestral (arboreal) and derived (bipedal) traits, which indicates a mixed locomotion and a more mosaic evolution of the trait. Some early hominins appear to, for instance, have bowl-shaped pelvises (hip bones) and angled femurs suitable for bipedalism but also have retained an opposable big toe or curved fingers and longer arms (for arboreal locomotion). These mixed morphologies may indicate that earlier hominins were not fully obligate bipeds and thus thrived in mosaic or mixed environments.
However, among Australopiths (members of the genus, Australopithecus discussed below), there are indications for greater reliance on bipedalism as the primary form of locomotion. Similarly, adaptations consistent with tool manufacture (shorter fingers and a longer, more robust thumb, in contrast to the features associated with arborealism) have been argued to appear before the genus Homo.
Several ideas about why bipedalism, which involves a radical restructuring of the body, was favored. Some think the shift to bipedalism is a response to an episode of drying in Africa, during which tropical rainforests developed into savanna/woodland environments. Bipedalism may have been a response to this environmental shift. Bipedalism as discussed, entails costs, such as stress on the lower spine and knees, especially for pregnant females. It must therefore have conferred some advantage given these drawbacks. Bipedalism may have helped hominins see further, and it could have helped hominins to carry objects, transport food, and make tools. Bipedalism may also have enabled hominins to cool off better in more open environments. This enhanced thermoregulation stems from less surface area being exposed to direct sunlight in the heat of midday. Finally, bipedalism is more efficient than knuckle-walking for traveling long distances, potentially beneficial in a savanna/woodland environment. These benefits are not mutually exclusive and each may have played a role in selection for upright posture. As it stands, we know that early hominins were bipedal but are uncertain of the conditions under which this trait was selected for.
Table. 5.1. “Early Hominins” chapter in Explorations: An Open Invitation to Biological Anthropology,
|
Region |
Feature |
Obligate Biped (H. sapiens) |
Nonobligate Biped |
|
Cranium |
Position of the foramen magnum |
Positioned inferiorly (immediately under the cranium) so that the head rests on top of the vertebral column for balance and support (head is perpendicular to the ground). |
Posteriorly positioned (to the back of the cranium). Head is positioned parallel to the ground. |
|
Postthermoregulation cranium |
Body proportions |
Shorter upper limb (not used for locomotion). |
Longer upper limbs (used for locomotion). |
|
Post cranium |
Spinal curvature |
S-curve due to pressure exerted on the spine from bipedalism (lumbar lordosis). |
C-curve. |
|
Post cranium |
Vertebrae |
Robust lumbar (lower-back) vertebrae (for shock absorbance and weight bearing). Lower back is more flexible than that of apes as the hips and trunk swivel when walking (weight transmission). |
Gracile lumbar vertebrae compared to those of modern humans. |
|
Post cranium |
Pelvis |
Shorter, broader, bowl-shaped pelvis (for support); very robust. Broad sacrum with large sacroiliac joint surfaces. |
Longer, flatter, elongated ilia; more narrow and gracile; narrower sacrum; relatively smaller sacroiliac joint surface. |
|
Post cranium |
Lower limb |
In general, longer, more robust lower limbs and more stable, larger joints. Large femoral head and longer neck (absorbs more stress and increases the mechanical advantage). Valgus knee, in which the angle of the knee positions it over the ankle and keeps the center of gravity balanced over the stance leg during stride cycle (shock absorbance). Distal tibia (lower leg) of humans has a large medial malleolus for stability. |
In general, smaller, more gracile limbs with more flexible joints. Femoral neck is smaller in comparison to modern humans and shorter. The legs bow outward, and there is no valgus angle of the knee (no “knock knees”). The distal tibia in chimpanzees is trapezoid (wider anteriorly) for climbing and allows more flexibility. |
|
Post cranium |
Foot |
Rigid, robust foot, without a midtarsal break. Nonopposable and large, robust big toe (for push off while walking) and large heel for shock absorbance. |
Flexible foot, midtarsal break present (which allows primates to lift their heels independently from their feet), opposable big toe for grasping. |
Early Hominins: Sahelanthropus and Orrorin
The authors of “Early Hominins” chapter in Explorations: An Open Invitation to Biological Anthropology, discuss bipedalism in the earliest hominin species . We see evidence for bipedalism in some of the earliest fossil hominins, dated from within our estimates of our divergence from chimpanzees. These hominins, however, also indicate evidence for arboreal locomotion. The earliest dated hominin find (between 6 mya and 7 mya, based on radiometric dating of volcanic tufts) is named Sahelanthropus tchadensis (Figure 5.6; Brunet et al. 1995). The initial discovery was made in 2001 by Ahounta Djimdoumalbaye and announced in Nature in 2002 by a team led by French paleontologist Michel Brunet. The find has a small cranial capacity (360 cc) and smaller canines than those in extant great apes, though they are larger and pointier than those in humans. This implies strongly that, over evolutionary time, the need for display and dominance among males has reduced, as has our sexual dimorphism. A short cranial base and a foramen magnum that is more humanlike in positioning have been argued to indicate upright walking.
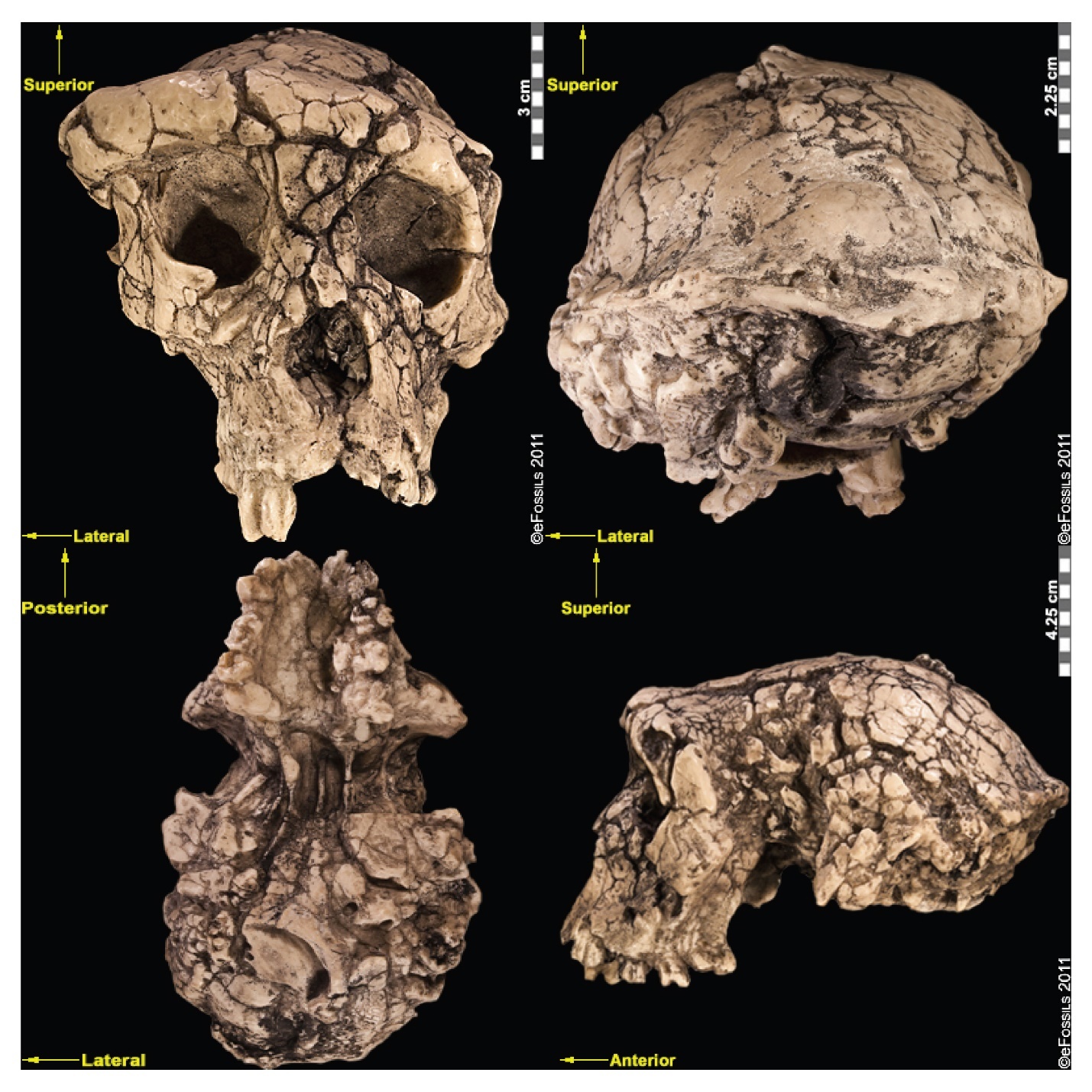
Initially, the inclusion of Sahelanthropus in the hominin family was debated by researchers, since the evidence for bipedalism is based on cranial evidence alone, which is not as convincing as postcranial evidence. Yet, a femur (thigh bone) and ulnae (upper arm bones) thought to belong to Sahelanthropus was discovered in 2001 (although not published until 2022). These bones may support the idea that the hominin was in fact a terrestrial biped with arboreal capabilities and behaviors (Daver et al. 2022).
Orrorin tugenensis (Orrorin meaning “original man”), dated to between 6 mya and 5.7 mya, was discovered near Tugen Hills in Kenya in 2000. Smaller cheek teeth (molars and premolars) than those in even more recent hominins, thick enamel, and reduced, but apelike, canines characterize this species. This is the first species that clearly indicates adaptations for bipedal locomotion, with fragmentary leg, arm, and finger bones having been found but few cranial remains. One of the most important elements discovered was a proximal femur, BAR 1002’00. The femur is the thigh bone, and the proximal part is that which articulates with the pelvis; this is very important for studying posture and locomotion. This femur indicates that Ororrin was bipedal, and recent studies suggest that it walked in a similar way to later Pliocene hominins. Some have argued that features of the finger bones suggest potential tool-making capabilities, although many researchers argue that these features are also consistent with climbing.
Early Hominins: The Genus Ardipithecus
Another genus, Ardipithecus, is argued to be represented by at least two species: Ardipithecus (Ar.) ramidus and Ar. kadabba.
Ardipithecus ramidus (“ramid” means root in the Afar language) is currently the best-known of the earliest hominins (Figure 9.9). Unlike Sahelanthropus and Orrorin, this species has a large sample size of over 110 specimens from Aramis alone. Dated to 4.4 mya, Ar. ramidus was found in Ethiopia (in the Middle Awash region and in Gona). This species was announced in 1994 by American paleoanthropologist Tim White, based on a partial female skeleton nicknamed “Ardi” (ARA-VP-6/500; White et al. 1994). Ardi demonstrates a mosaic of ancestral and derived characteristics in the postcrania. For instance, she had an opposable big toe (hallux), similar to chimpanzees (i.e., more ancestral), which could have aided in climbing trees effectively. However, the pelvis and hip show that she could walk upright (i.e., it is derived), supporting her hominin status. A small brain (300 cc to 350 cc), midfacial projection, and slight prognathism show retained ancestral cranial features, but the cheek bones are less flared and robust than in later hominins.
Exercises
Australopithecines
Some of the earliest and best understood hominins were Australopithecines, all of which have so far been discovered in Africa. There are several different species of australopithecines: Australopithicus afarensis, Australopithecus africanus, Australopithecus sebida and several others. Skeletal morphology (the form of the skeleton) reveals that Australopithecines walked on two legs as their main means of locomotion (moving around). No other primate besides humans is habitually bipedal.
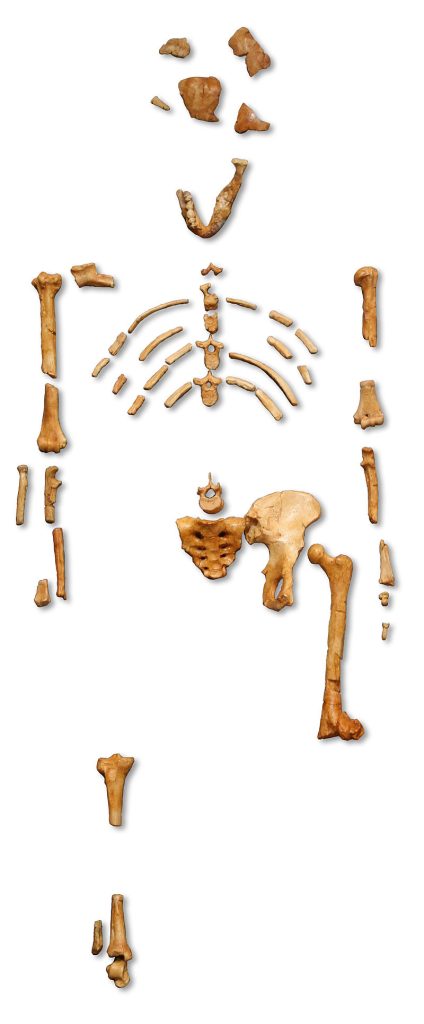
The Hips Don’t Lie: Evidence for Bipedalism
The most famous Australopithecine is probably “Lucy”, an Australopithecus afarensis, discovered by Donald Johanson in Hadar, Ethiopia, in 1974. Lucy dates to about 3–3.5 million years ago (mya). She is remarkable not so much for her age, but for the completeness of her skeleton, with 40% of the bones being present. Lucy was about 20 years old and just under 4 feet when she died. Her brain was small (chimp-sized), but she was clearly bipedal. Much of her postcranial skeleton (from the neck down) was present. Her pelvis, or hip bones, were especially informative. Lucy’s hip structure was more “bowl-shaped” like humans, and her femurs and knee joints angled inward, as in humans, to maintain a central center of gravity while walking upright. In contrast, chimpanzees have an elongated pelvis and femurs (upper leg bone) and knees that do not angle in. Therefore, the structure of the Australopithecine hip indicates bipedalism.
Sarah S. King, Kara Jones, and Lee Anne Zajicek describe an even more complete Australopithecine find in “Chapter 7: Understanding the Fossil Context” in Explorations: An Open Invitation to Biological Anthropology. In 1994, an Australopithecus fossil nicknamed “Little Foot” (Stw 573) was located in the World Heritage Site at Sterkfontein Caves (“the Cradle of Humankind”) in South Africa. Little Foot is more complete than Lucy and possibly the oldest fossil that has so far been found, dating to at least 3.6 million years (Granger et al. 2015). The ankle bones of the fossil were extricated from the matrix of concrete-like rock, revealing that the bones of the ankles and feet indicate bipedalism (University of Witwatersrand 2017).
Both the Lucy and Little Foot fossils date back to the Pliocene (5.8 to 2.3 mya). Older hominin fossils from the late Miocene (7.25 to 5.5 mya) have been located, although they are much less complete. The oldest hominin fossil is a fragmentary skull named Sahelanthropus tchadensis, found in Northern Chad and dating to circa seven mya (Lebatard et al. 2008). It is through the discovery, dating, and study of primate and early hominin fossils that we find physical evidence of the evolutionary timeline of humans.
Other indications of bipedalism in Australopithecines include the absence of a divergent big toe, as chimps have. The australopithecine toe is in line with the others making it adaptive to bipedal walking. The foramen magnum, which is the large opening for the spinal cord at the base of skull, is oriented downward, indicating the head sat atop the spinal column. Based on the arm bones, however, Australopithecines may have been partially arboreal.

Another piece of evidence for bipedalism in Australopithecines is the famed Laetoli footprints in Tanzania, discovered in 1978 (Figure 5.8). At Laetoli, numerous animal prints have been preserved in volcanic ash. Following a volcanic eruption, hominins (and other creatures) walked on ash, creating footprints, which were subsequently buried by more ash and preserved. The hominin footprint showed no evidence of quadrupedalism or knuckle walking and were dated to around 3.6 million years ago. In all, 70 hominin footprints created by three individuals were discovered. In 2016, two new trackways at Laetoli were reported (Ichumbaki et al. 2016). Computer modeling of the footprints reveals that Australopithecines walked much like we do today in terms of weight transfer and push-off. (Crompton 2012)
Defining the Genus Homo
Bonnie Yoshida-Levine explains the criteria typically used to define our genus, the genus Homo in “Early Members of the Genus Homo” in Explorations and Open Invitation to Biological Anthropology, second edition. Because Anthropology is fundamentally concerned with what makes us human, defining our own genus takes on special significance for anthropologists. Ever since scientists acknowledged the existence of extinct species of humans, they have debated which of them display sufficient “humanness” to merit classification in the genus Homo. When grouping species into a common genus, biologists consider criteria such as physical characteristics (morphology), evidence of recent common ancestry, and adaptive strategy (use of the environment). However, there is disagreement about which of those criteria should be prioritized, as well as how specific fossils should be interpreted in light of the criteria.
Nevertheless, there is general agreement that species classified as Homo should share characteristics that are broadly similar within our species. These include the following:
- a relatively large brain size, indicating a high degree of intelligence;
- a smaller and flatter face;
- smaller jaws and teeth; and
- increased reliance on culture, particularly the use of stone tools, to exploit a greater diversity of environments (adaptive zone).
Some researchers would include larger overall body size and limb proportions (longer legs/shorter arms) in this list. While these criteria seem relatively clear-cut, evaluating them in the fossil record has proved more difficult, particularly for the earliest members of the genus. There are several reasons for this. First, many fossil specimens dating to this time period are incomplete and poorly preserved. Second, early Homo fossils appear quite variable in brain size, facial features, and teeth and body size, and there is not yet consensus about how to best make sense of this diversity. Finally, there is growing evidence that the evolution of the genus Homo proceeded in a mosaic pattern: in other words, these characteristics did not appear all at once in a single species; rather, they were patchily distributed in different species from different regions and time periods. Consequently, different researchers have come up with conflicting classification schemes depending on which criteria they think are most important.
Homo habilis in Middle Earth
The first hominin that is sometimes classified into our own genus—the genus Homo—evolved in eastern Africa from Australopithecine populations around 2.1–1.5 million years ago and is called Homo habilis. The term Homo habilis roughly means “handy man” in Latin. Compared to australopithecines, habilis has more human-like features including the cranial capacity is larger (about 650 cc compared to 500cc) and it has a less prognathic (projecting forward) face and a more vertical forehead.
Habilis is thought to be one of the first hominins to make and use stone tools—the Oldowan stone tool tradition, though stone tool cut marks are known from a pre-habilis site (Lovett 2010). Captive chimps have been shown by human trainers how to smash up long bones and use the splinters to puncture containers full of Kool-Aid. Monkeys produce flakes from smashing rocks, but so far, they haven’t used them to cut with. Making and using sharp edges seems to be within the performance capabilities of apes and even monkeys, but applying cutting edge technology in the wild is not in the repertoire. This propensity for making stone tools, is quite a bit more like modern humans than apes. It seems likely that cutting edge production in hominins was conditioned or selected for by a distinctive subsistence pattern, namely a strong dependence on hunting and/or scavenging meat. It is thought that Homo habilis scavenged for meat.
Habilis was fully bipedal, although it had not yet developed the modern human body proportions; it lacks the longer legs and shorter torso adapted for running and striding, that we see in later hominins. There are relatively few postcranial fossils from Homo habilis and different specimens show a range of variation, making it a controversial species. Many researchers lump them together with Australopithecines, considering them all to be a continuous, highly variable species. One paleoanthropologist has suggested that the world of early Homo was like Middle Earth in The Hobbit, inhabited by many very different looking bipedal primates.
Yoshida-Levine points out controversies over Homo habilis in “Early Members of the Genus Homo.” Since its initial discovery, many more Homo habilis were discovered in East and South African sites during the 1970s and 1980s. As more fossils joined the ranks of Homo habilis, several trends became apparent. First, the fossils were quite variable. While some resembled the fossil specimen first published by Leakey and colleagues, others had larger cranial capacity and tooth size. A well-preserved fossil skull from East Lake Turkana labeled KNM-ER-1470 displayed a larger cranial size along with a strikingly wide face. The diversity of the Homo habilis fossils prompted some scientists to question whether they displayed too much variation to all belong to the same species. They proposed splitting the fossils into at least two groups. The first group resembling the original small-brained specimen would retain the species name Homo habilis; the second group consisting of the larger-brained fossils such as KNM-ER-1470 would be assigned the new name of Homo rudolfensis (see Figure 5.10). Researchers who favored keeping all fossils in Homo habilis argued that sexual dimorphism, adaptation to local environments, or developmental plasticity could be the cause of the differences. For example, modern human body size and body proportions are influenced by variations in climates and nutritional circumstances.
Yoshida-Levine points out in “Early Members of the Genus Homo” that the known dates for fossils identified as Homo habilis range from about 2.5 million years ago to 1.7 million years ago. Recently, a partial lower jaw dated to 2.8 million years from the site of Ledi-Gararu in Ethiopia has been tentatively identified as belonging to the genus Homo (Villmoare et al. 2015). If this classification holds up, it would push the origins of our genus back even further.
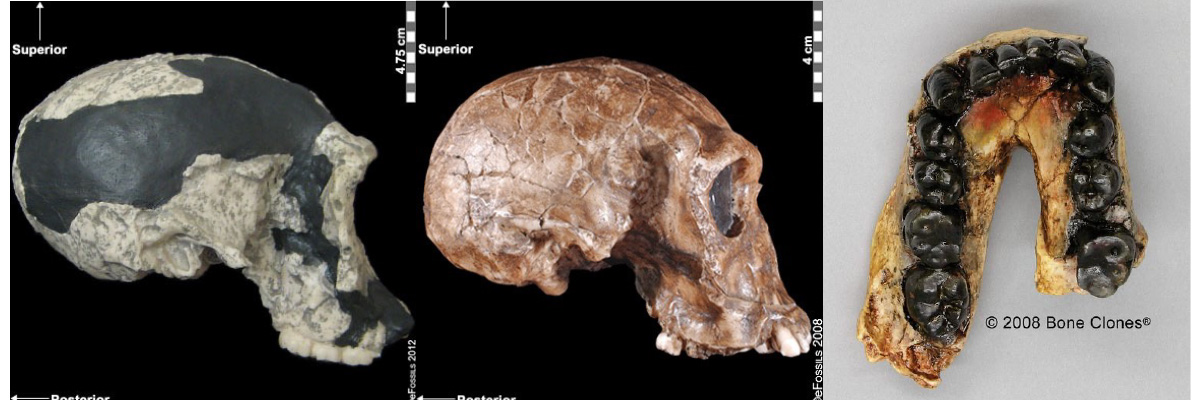
Figure 5.10. Homo habilis fossil specimens. From left to right they are: a. lateral right view of OH-24 (found at Olduvai Gorge), b. lateral right view of KNM-ER-1813 (from Koobi Fora, Kenya), and c. the jaw of OH-7, which was the type specimen found in 1960 at Olduvai Gorge, Tanzania. Credit: a. Homo habilis: OH 24 lateral right view by ©eFossils is under a CC BY-NC-SA 2.0 License and is used as outlined by eFossils. b. Homo habilis: KNM-ER 1813 lateral right view by ©eFossils is under a CC BY-NC-SA 2.0 License and is used as outlined by eFossils. c. Homo habilis OH 7 Jaw by ©BoneClones is used by permission and available here under a CC BY-NC 4.0 License.
Early Stone Tools
Yoshida-Levine writes in “Early Members of the Genus Homo,” that the larger brains and smaller teeth of early Homo are linked to a different adaptive strategy than that of earlier hominins: one dependent on modifying rocks to make stone tools and exploit new food sources. Stone tools become more frequent at sites dating to about 2 million years ago, the time of Homo habilis (Roche et al. 2009). This suggests that these hominins were increasingly reliant on stone tools to make a living.
Stone tools are assigned a good deal of importance in the study of human origins. Examining the form of the tools, the raw materials selected, and how they were made and used can provide insight into the thought processes of early humans and how they modified their environment in order to survive. Paleoanthropologists have traditionally classified collections of stone tools into industries, based on their form and mode of manufacture. There is not an exact correspondence between a tool industry and a hominin species; however, some general associations can be made between tool industries and particular hominins, locations, and time periods.
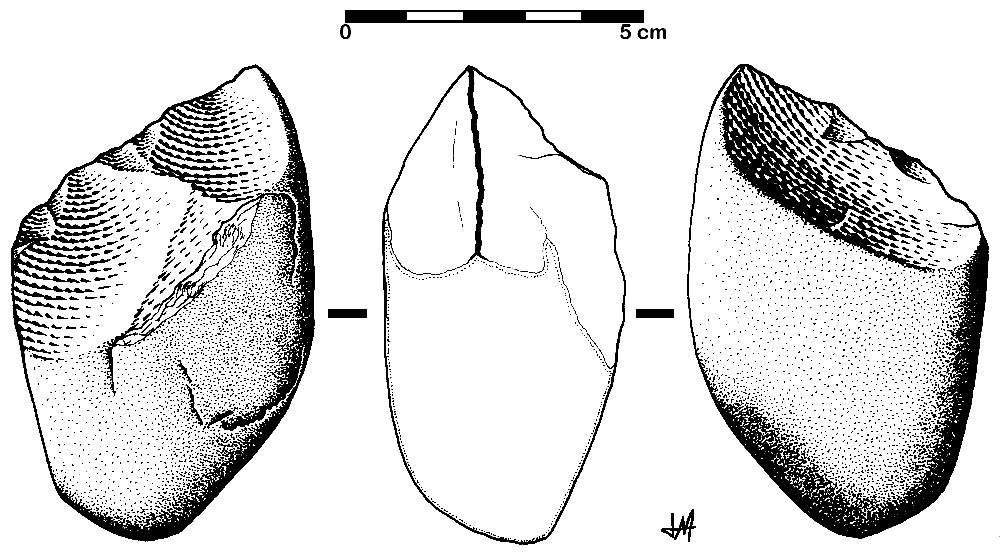
The Oldowan tool industry is named after the site of Olduvai Gorge in Tanzania where the tools were first discovered. The time period of the Oldowan is generally estimated to be 2.5 mya to 1.6 mya. The tools of this industry are described as “flake and chopper” tools—the choppers consisting of stone cobbles with a few flakes struck off them (Figure 5.11). To a casual observer, these tools might not look much different from randomly broken rocks. However, they are harder to make than their crude appearance suggests. The rock selected as the core must be struck by the rock serving as a hammerstone at just the right angle so that one or more flat flakes are removed. This requires selecting rocks that will fracture predictably instead of chunking, as well as the ability to plan ahead and envision the steps needed to create the finished product. The process leaves both the core and the flakes with sharp cutting edges that can be used for a variety of purposes.
{kind=link}
Stone Tool Use and the Diet of Early Homo
Yoshida-Levine asks in “Early Members of the Genus Homo,” What were the hominins doing with the tools? One key activity seems to have been butchering animals. Studies of animal bones at the site show cut marks on bones, and leg bones are often cracked open, suggesting that they were extracting the marrow from the bone cavities. It is interesting to consider whether the hominins hunted these animals or acquired them through other means. The butchered bones come from a variety of African mammals, ranging from small antelope to animals as big as wildebeest and elephants! It is difficult to envision slow, small-bodied Homo habilis with their Oldowan tools bringing down such large animals. One possibility is that the hominins were scavenging carcasses from lions and other large cats. Paleoanthropologist Robert Blumenschine has investigated this hypothesis by observing the behavior of present-day animal carnivores and scavengers on the African savanna. When lions abandon a kill after eating their fill, scavenging animals arrive almost immediately to pick apart the carcass. By the time slow-footed hominins arrived on the scene, the carcass would be mostly stripped of meat. However, if hominins could use stone tools to break into the leg bone cavities, they could get to the marrow, a fatty, calorie-dense source of protein (Blumenschine et al. 1987). Reconstructing activities that happened millions of years ago is obviously a difficult undertaking, and paleoanthropologists continue to debate whether scavenging or hunting was more commonly practiced during this time.
Regardless of how they were acquiring the meat, these activities suggest an important dietary shift from the way that the australopithecines were eating. The Oldowan toolmakers were exploiting a new ecological niche that provided them with more protein and calories. And it was not just limited to meat-eating—stone tool use could have made available numerous other subsistence opportunities. A study of microscopic wear patterns on a sample of Oldowan tools indicates that they were used for processing plant materials such as wood, roots or tubers, and grass seeds and stems (Lemorini et al. 2014). In fact, it has been pointed out that the Oldowan toolmakers’ cutting ability (whether for the purposes of consuming meat and plants or for making tools, shelters, or clothing) represents a new and unique innovation, never seen before in the natural world (Roche et al. 2009).
Homo erectus
By about 2.0 to 1.8 mya, Homo erectus represents a continuation of anatomical and behavioral trends already present in Homo habilis—increased encephalization, stone tool use, reduced prognathism and teeth, and of course bipedalism. Erectus had a larger relative brain size than habilis, but still smaller than modern humans. Erectus also had a thick skull and massive browridge that continued, shelf-like, across the forehead.
Yoshida-Levine summarizes differences between Homo habilis and Homo erectus in “Early Members of the Genus Homo.” Compared to Homo habilis, Homo erectus showed increased brain size, smaller teeth, and a larger body. However, it also displayed key differences from later hominin species including our own. Although the head of Homo erectus was less ape-like in appearance than the australopithecines, it did not resemble modern humans (Figure 5.12). Compared to Homo habilis, Homo erectus had a larger brain size: an average of about 900 cc compared to 650 cc to 750 cc. Instead of a rounded shape like our skulls, the erectus skull was long and low like a football, with a receding forehead, and a horizontal ridge called an occipital torus that gave the back of the skull a squared-off appearance. The cranial bones are thicker than those of modern humans, and some Homo erectus skulls have a slight thickening along the sagittal suture called a sagittal keel. Large, shelf-like brow ridges hang over the eyes. The face shows less prognathism, and the back teeth are smaller than those of Homo habilis. Instead of a pointed chin, like ours, the mandible of Homo erectus recedes back.
The biggest change was in the development of its nearly modern postcranial skeleton (that is, body proportions and stature). This is thought to be associated with an adaptation for striding, running and covering large distances during the day while foraging, probably indicating an increased dependence on hunting animals. Yoshida-Levine writes that unlike Homo habilis and the australopithecines, both of whom were small-statured with long arms and short legs, Homo erectus shows evidence of being fully committed to life on the ground. This meant long, powerfully muscled legs that enabled these hominins to cover more ground efficiently. Indeed, studies of the Homo erectus body form have linked several characteristics of the species to long-distance running in the more open savanna environment (Bramble and Lieberman 2004). Erectus is the first hominin to be found outside of Africa. There is variability between erectus fossils, especially between Asia and Africa, and some designate the African erectus as Homo ergaster.

Our best evidence for reconstructing Homo erectus body build for comes from a fossil known as Turkana boy (or Nariokotome boy) *recovered from near the shore of Lake Turkana in Kenya, which dates to about 1.5 mya (Note: some researchers consider Turkana boy to be Homo ergaster, the African contemporary of Homo erectus). Turkana Boy is a near complete skeleton of a young boy, who based on tooth eruption patterns was about 10 or 11 years old at the time of his death. He stood about 5 feet tall when he died, and would have grown to about 5’ 6” as an adult. It is not known what caused his death or how he came to be so quickly buried by sediments and was thereby so incredibly well preserved. Despite his modern body build and stature, his cranial capacity as an adult would have been about 900 cc, about 440 cc less than the average brain size of modern people.
By comparing tibia (the larger lower leg bone) lengths of adult erectus specimens, paleoanthropologists have estimated the average height of Homo erectus to be between 5’5 and 5’7”. This is close to the average height of modern Europeans in the 18th and 19th century. Furthermore, this average height stays stable for about 2 million years, through the appearance of later species. Anatomically modern humans, that is, humans identical in morphology to us, who appeared about 200-300 thousand years ago in Africa, were slightly taller, averaging about 5’10”, which is taller than the average height of most present-day people. The take-home message here is that our modern body proportions and stature evolved early—much earlier than our modern brain size—and has stayed constant ever since. This suggests that this body build was a fundamental aspect of our adaptation to the environment.
Homo erectus emerged during the geological time period marked by a series of ice ages and radical fluctuation in the Earth’s climate. This time period, called the Pleistocene began around 2.5 million years ago and ended around 12,000 years ago. The Pleistocene consisted of a series of glacial periods, periods of long-term reduction in the temperature of the Earth’s surface and atmosphere, resulting in the presence or expansion of continental and polar ice sheets and alpine glaciers, interrupted periodically by warmer interglacials. This cold-warm pattern alternated on a fairly regular cycle about 100,000 years long (see the Milankovitch cycles). The world’s configuration of continents was the same as today, but some land masses were exposed because much of the world’s water was bound up in enormous ice sheets, more than 2 miles thick in places. Ocean levels at one point were 330 feet lower than present levels.
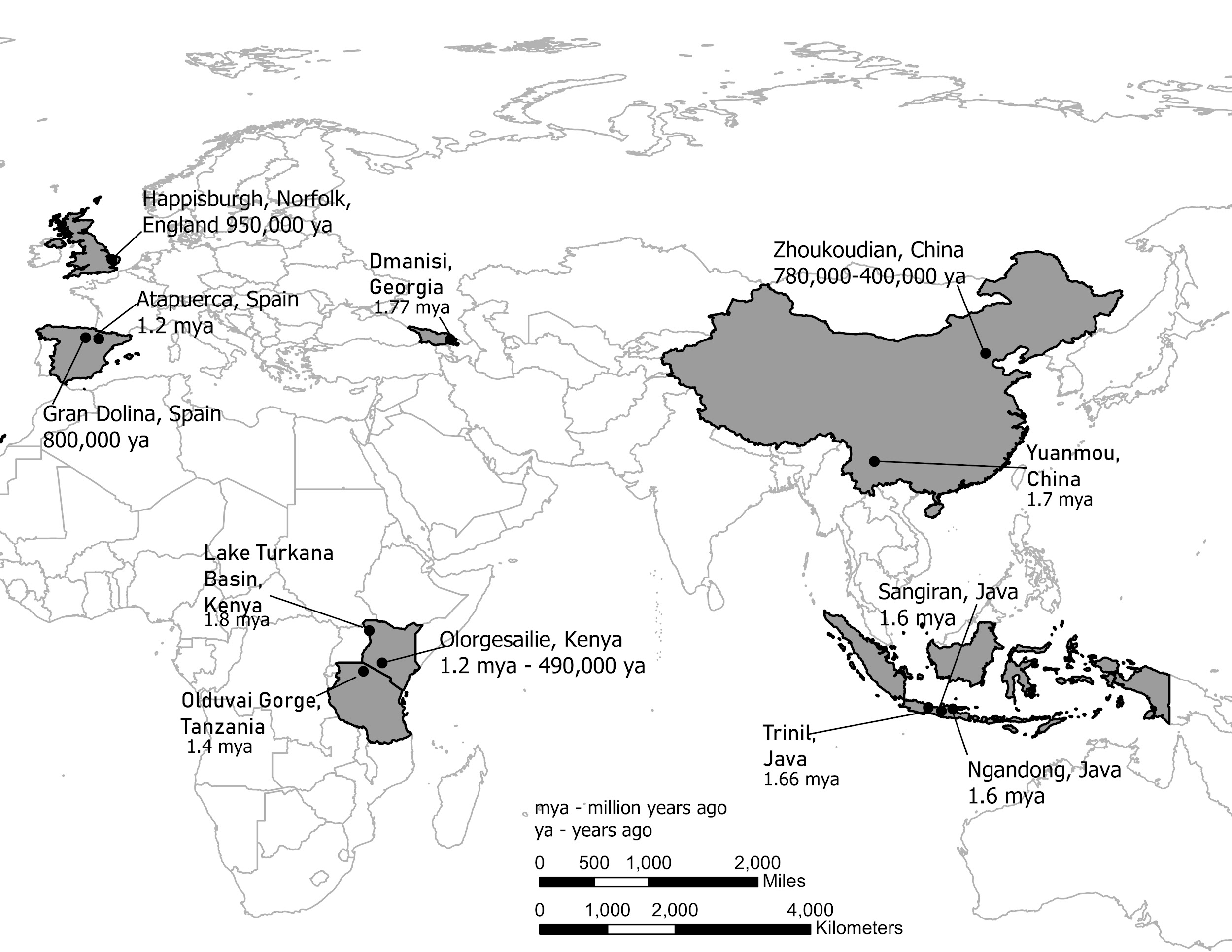
It is generally agreed that Homo erectus was the first hominin to migrate out of Africa and colonize Asia and later Europe (although recent discoveries in Asia may challenge this view, Figure 5.14). Unlike previous species, fossils and archaeological sites of Homo erectus are found not only in Africa, but also in Europe and Asia. Some erectus fossils have been found on islands in southeast Asia, like Java, which means these hominins could make rafts or boats, perhaps out of the bamboo that grows throughout the region, to cross significant expanses of open sea. They were also able to survive and prevail in temperate climates with extended freezing winter weather in places like England, northern Europe, and northern China, which suggests that they could make and control fire for warmth.
A site in Dmanisi, Republic of Georgia suggests the picture of Homo erectus may be more complex than previously thought (Figure 5.13). Georgia is a small country between Russia and Iran on the east coast of the Black Sea. Here, in Pleistocene fossil deposits, five well-preserved Homo erectus crania were discovered around 2010 (Skull 3). The Homo erectus skulls at Dmanisi are relatively small in size. One of those skulls (Skull 5) looks very primitive, meaning more like earlier fossils, with a very small braincase (346 cc) and prominent canines, more like Homo habilis than erectus. What’s more, the associated stone tools are of the earlier Oldowan industry, not the more developed Acheulean industry typical of Homo erectus. The tools at Dmanisi were associated with butchered bones of large grazing animals typical of a savannah, or grassland environment. The site dates to as old as 1.8 mya. The discoveries at Dmanisi mean that genus Homo ventured out of Africa much earlier than previously recognized, and at a much more primitive level of development with respect to body build, cranial capacity and tool kit.

Figure 5.13: “The site of Dmanisi” by Larry Dumlao is licensed under CC BY-SA 4.0.
The variation at Dmanisi is surprising. Had the fossils skulls been found at different sites, they may have been designated as different species (SciNews 2013). This points to an ongoing problem in assigning fossils to taxonomic categories, namely, it is hard to know whether a fossil represents a single species or variation within a single species.
Handaxes

The appearance of Homo erectus coincides with a new, more sophisticated stone tool type called the Acheulean handaxe (Figure 5.15, 5.16). These tools could be used for a variety of purposes, like digging or cutting. In addition, they could be used to produce sharp flakes of stone that were tools in and of themselves. Acheulian handaxes were widespread and took a sophisticated brain to produce. Kanzi, the bonobo chimpanzee, can produce a simple flake to cut a rope, but no chimpanzee in captivity or in the wild has made anything close to an Acheulian handaxe.

In reconstructing the diet of Homo erectus, researchers can draw from multiple lines of evidence. These include stone tools used by Homo erectus, animal bones and occasionally plant remains from Homo erectus sites, and the bones and teeth of the fossils themselves. These data sources suggest that compared to the australopithecines, Homo erectus consumed more animal protein. Coinciding with the appearance of Homo erectus fossils in Africa are archaeological sites with much more abundant stone tools and larger concentrations of butchered animal bones (Figure 5.17).

Yoshida-Levine writes that, it makes sense that a larger body and brain would be correlated with a dietary shift to more calorically dense foods. This is because the brain is a very energetically greedy organ. Indeed, our own human brains require more than 20% of one’s calorie total intake to maintain. When biologists consider the evolution of intelligence in any animal species, it is often framed as a cost/benefit analysis: For large brains to evolve, there has to be a compelling benefit to having them and a way to generate enough energy to fuel them.
Exercises
Attribution
This chapter is a mixture of the following sources.
Kerryn Warren, Lindsay Hunter, Navashni Naidoo, Silindokuhle Mavuso, “Chapter 9: Early Hominins” by Kerryn Warren, K. Lindsay Hunter, Navashni Naidoo, Silindokuhle Mavuso, Kimberleigh Tommy, Rosa Moll, and Nomawethu Hlazo. In Explorations: An Open Invitation to Biological Anthropology, second edition, edited by Beth Shook, Katie Nelson, Kelsie Aguilera, and Lara Braff, which is licensed under CC BY-NC 4.0. (The second edition of the chapter is a revision of “Chapter 9: Early Hominins” by Kerryn Warren, K. Lindsay Hunter, Navashni Naidoo, Silindokuhle Mavuso, Kimberleigh Tommy, Rosa Moll, and Nomawethu Hlazo. In Explorations: An Open Invitation to Biological Anthropology, first edition, edited by Beth Shook, Katie Nelson, Kelsie Aguilera, and Lara Braff, which is licensed under CC BY-NC 4.0.)
Bonnie Yoshida-Levine. “Chapter 10: Early Members of the Genus Homo” In Explorations: An Open Invitation to Biological Anthropology, second edition, edited by Beth Shook, Katie Nelson, Kelsie Aguilera, and Lara Braff, which is licensed under CC BY-NC 4.0. The second edition chapter is a revision from “Chapter 10: Early Members of the Genus Homo” by Bonnie Yoshida-Levine. In Explorations: An Open Invitation to Biological Anthropology, first edition, edited by Beth Shook, Katie Nelson, Kelsie Aguilera, and Lara Braff, which is licensed under CC BY-NC 4.0.)
Susan Ruth, “Chapter 9: How Did We Get Here?” Being Human, which is licensed under CC BY-NC 4.0.
Media Attributions
- 4917203005_050b785f2d_w
The locomotor ability to walk on two legs.
Species that are regarded as human, directly ancestral to humans, or very closely related to humans.
The ancient organism from which all living things on Earth are descended.
Where the primary form of locomotion for an organism is bipedal.
paleoanthropological hoax where a human skull fragment and an orangutan jaw were presented as a single fossil, claiming to be a missing link between apes and humans.
A trait that has been recently modified, most helpful when assigning taxonomic classification.
The angle of the knee between the femur and tibia, which allows for weight distribution to be angled closer to the point above the center of gravity (i.e., between the feet) in bipeds.
The big toe.
Different characteristics evolve at different rates and appear at different stages.
Maintaining body temperature through physiologically cooling or warming the body.
Or hind dentition (molars and premolars).
Closer to the main part of the body or the point of attachment, while "distal" means further away from that point.
Geological epoch dating from 5.3 to 2.6 million years ago.
The capability of an organism to modify its phenotype during development in response to environmental cues.
Earliest stone-tool industry consisting of simple flakes and choppers.
A ridge on the occipital bone in the back of the skull.
A thickened area along the top of the skull.
Condition where the lower face and jaw protrude forward from a vertical plane.